Glass sponges occur worldwide, mostly at depths between 200 and 1000 m. This group of sponges are especially abundant in the Antarctic.
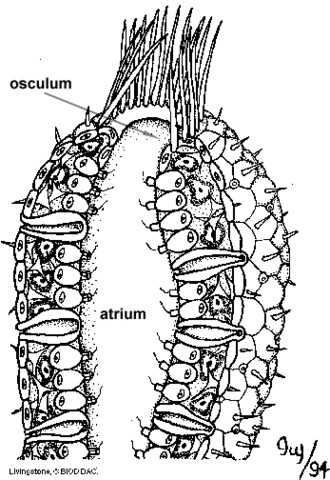
All glass sponges are upright, and possess specialized structures at their bases for holding fast to the ocean floor. Most appear outwardly to be radially symmetrical; they are typically cylindrical, but may also be cup-shaped, urn-shaped, or branching. The average height of a hexactinellid is between 10 and 30 cm, but some can grow to be quite large. A hexactinellid possesses a cavernous central cavity (the atrium ) through which water passes; a cap of tightly woven spicules covers the osculum in some species. Coloration in most is pale. Glass sponges most closely resemble syconoid sponges, but they differ too much internally from other sponges to be considered truly syconoid.
{kind=link}
It is upon close internal examination that glass sponges can be most easily distinguished from other sponges. The skeleton of an hexactinellid is made entirely of silica. These siliceous spicules are generally composed of three perpendicular rays (and therefore six points, so they are described as hexactine), and are often fused, lending hexactinellids a structural rigidity not typical of other sponge taxa. Strung between the spicules is a largely syncytial network of soft body cells. Incurrent water enters the body through spaces in the syncytial strands . Within the syncytia are units functionally similar to the choanocytes found in other sponges but these units completely lack nuclei, and so are referred to as collar bodies rather than collar cells. They are flagellated, and it is the beating of their flagella that causes the current to pass through the sponge. Within the syncytia are cells functionally comparable to archaeocytes in other sponges, but these cells seem to demonstrate only limited mobility. Hexactinellids lack myocytes completely, and so are incapable of contraction. While hexactinellids possess no nerve structure, they seem to be able to send electrical signals across the body through the syncytial soft tissue.
{kind=link}
Little is known about hexactinellid reproduction and development. Sperm are taken into an organism with water, and then must make their way to eggs within the organism. After fertilization, the larvae are incubated for a relatively long time so they even form rudimentary spicules before being released as parenchymella larvae. These differ from other sponge larvae in lacking flagella or any other method of locomotion. Hexactinellids cluster to an unusually high degree, suggesting that larvae do not drift far before settling. After a larva lands on the ocean floor, it metamorphoses, and the adult sponge begins to grow. Hexactinellids are known for prolific budding.
Glass sponges are purely filter feeders. Sponges subsist on macroscopic detritus material, but also consume cellular material, bacteria, and nonliving particles so small they cannot be resolved with a light microscope. Small particles of edible material taken in by the current created by collar bodies are absorbed as they pass through the channels within the sponge. The collar bodies are covered with microvilli that trap food, and the food passes through vacuoles through the collar bodes and into the syncytia. Archaeocytes between the syncytial strands are responsible for food distribution and storage. The archaeocytes may also be responsible to some extent for food capture. Hexactinellids seem to lack selective control over the food they ingest - any food small enough to penetrate the syncytium is ingested. Because of their lack of a continuous outer membrane and their lack of defined ostia, hexactinellids lack control over how much water passes through them. It is believed that the stability of deep-water environments allows hexactinellids to survive despite these shortcomings.
Hexactinellids are completely sessile. Even larvae seem to display no movement, outside of their ability to disperse small distances in currents. Furthermore, unlike other sponges, hexactinellids do not contract when stimulated.
As with other sponges, hexactinellids may be sources of pharmaceuticals, although their economic potential is largely unexploited. Humans rarely come into contact with glass sponges, and are mostly unaffected by them. In Japan, however, they are given as wedding presents. Hexactinellids of a particular species engage in a symbiotic relationship with shrimp. When small, two shrimp of opposite sexes enter the sponge atrium, and, after growing to a certain size, cannot leave. They feed on material brought in by the currents produced by the sponge, and eventually reproduce. A skeleton of a sponge containing the two shrimp is given as a wedding present in Japan.
Little effort is being made to preserve hexactinellid species, but there may be great value in keeping glass sponge populations healthy, as they may hold the secrets of hundreds of millions of years of evolution, and may have evolved chemicals of potential value to humanity.
Hexactinellids are considered close relatives of Demospongiae .
References:
Ax. 1996. Multicellular Animals: A New Approach to the Phylogenetic Order in Nature; Springer, Berlin.
Bergquist, P. R. 1978. Sponges. University of California Press, Berkeley and Los Angeles. 268 pages.
Dohrmann, M., J. Dorte, J. Reitner, A.G. Collins, and G. Wörheide. 2008. Phylogeny and Evolution of Glass Sponges (Porifera, Hexactinellida). Systematic Biology, 57:388.
Kozloff, E. N. 1990. Invertebrates. Saunders College Publishing, Philadelphia and other cities. 866 pages.
Levin, H. L. 1999. Ancient Invertebrates and Their Living Relatives. Prentice Hall, Upper Saddle River. 358 pages.
Leys, S.P., Mackie, G.O., Reiswig, H.M. 2007. The biology of glass sponges. Advances in Marine Biology. 52:1¬145.
Philippe et al. 2009. Phylogenomics revives traditional views on deep animal relationships. Current Biology, 19:706.
Ruppert, E. E. and R. D. Barnes. 1994. Invertebrate Zoology: Sixth Edition. Saunders College Publishing. Fort Worth and other cities. 1040 pages.
Sperling, E.A., J.M. Robinson, D. Pisani, and K.J. Peterson. 2010. Where's the glass? Biomarkers, molecular clocks, and microRNAs suggest a 200-Myr missing Precambrian fossil record of siliceous sponge spicules. Geobiology 8:24.
Thiel, V., M. Blumenberg, J. Hefter, T. Paper, S. Pomponi, J. Reed, J. Reitner, G. Wörheide, and W. Michaelis. 2002. A chemical view of the most ancient metazoa--biomarker chemotaxonomy of hexactinellid sponges. Naturwissenschaften 89:60.
Additional Links
Contributors
Dan Atwater (author), Daphne G. Fautin (author).
- sessile
-
non-motile; permanently attached at the base.
Attached to substratum and moving little or not at all. Synapomorphy of the Anthozoa