Diversity
Platacanthomyidae is a small family of rodents. It contains just three species in
two genera:
Platacanthomys
, the spiny dormouse, and
Typhlomys
, the pygmy dormice.
Geographic Range
Platacanthomyids have a discontinuous Old World distribution. They are found in several
regions of southern India, southern China, and northern Vietnam.
- Biogeographic Regions
- palearctic
- oriental
Habitat
Platycanthomyids live in moist, rocky, tropical and subtropical forests at elevations
of 600 to 2100 meters. They inhabit burrows, tree cavities, and clefts between rocks,
often near streams.
- Habitat Regions
- tropical
- terrestrial
- Terrestrial Biomes
- forest
Physical Description
EXTERNAL CHARACTERISTICS:
Platacanthomyids are mouselike in overall appearance, with tail length ranging from 75 to 138 mm and total body length ranging from 70 to 212 mm. Long, stiff hairs form a brush on the tip of the tail. The feet are slim and small with medium long digits. Four of the digits on the front foot have claws, and the fifth is a rudimentary thumb with a nail. The soles of all four feet are naked and have six pads. Long vibrissae protrude from the relatively short muzzle. The ears are prominent and sparsely furred. Sexual dimorphism has not been described in this group.
DENTAL CHARACTERISTICS:
The dental formula is 1/1, 0/0, 0/0, 3/3 = 16. The cheek teeth are high-crowned and parallel ridges of enamel run diagonally across the crowns. The enamel on the incisors is orange. The molars in the upper jaw each have three roots; those in the lower jaw have two. The first two molars are about the same size, and the third is about 2/3 the size of the other two.
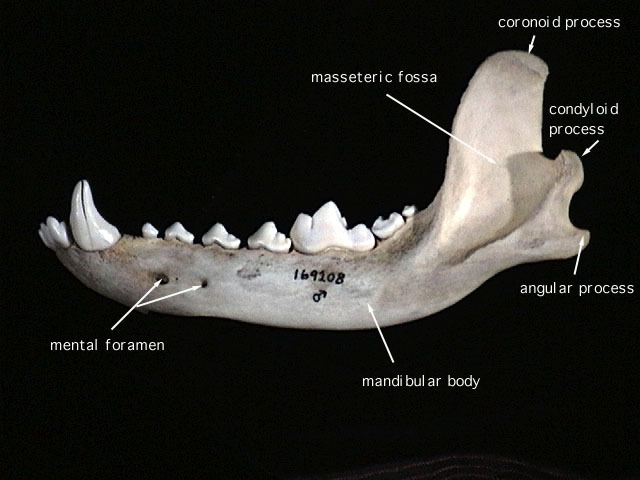
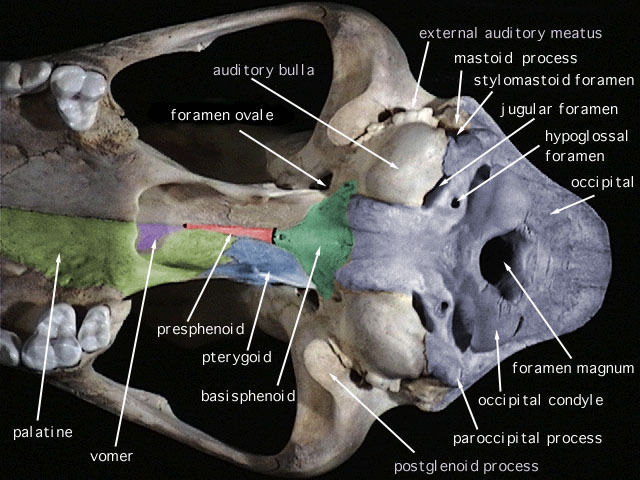
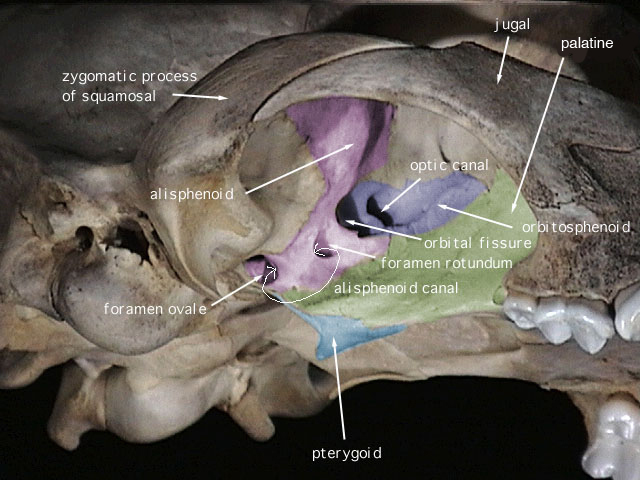
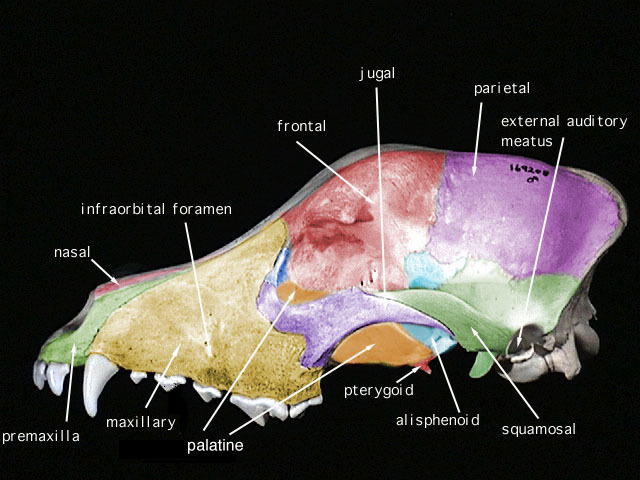
SKULL CHARACTERISTICS:
The small, delicate
dentary
has a low, angular
coronoid process
that in most specimens is positioned just slightly higher than the
condyloid process
. The unperforated
angular process
is not inflected lingually. The wide hard palate terminates anterior to the rear
margins of the molar rows. The interorbital region and the
interparietal
are both broad, the
occiput
is deep, and the
infraorbital foramina
are large and narrow. The lateral surface of the
alisphenoid canal
is formed by the
alisphenoid bone
. The
pterygoid
fossa, which may or may not be perforated with tiny holes, is broad, flat, and smoothly
continuous with the sides of the braincase. The masticatory-buccinator formanina
are coalesced into one opening. The complete, slightly enlarged mastoid is not perforated.
The small squamosomastoid foramen is contained within the suture between the
squamosal
and the mastoid. The
auditory bullae
are relatively small and lack transbullar septae.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
- Other Physical Features
- endothermic
- bilateral symmetry
Reproduction
No information is available on the mating system of platacanthomyids.
No information is available on the reproduction of platacanthomyids, besides the fact that they are eutherian mammals and therefore reproduce sexually via internal fertilization and bear live young.
- Key Reproductive Features
- gonochoric/gonochoristic/dioecious (sexes separate)
- sexual
- fertilization
- viviparous
Female platacanthomyids nurse their young, being mammals, but no other information is available on the investment that they make in their offspring.
- Parental Investment
-
pre-fertilization
- provisioning
-
protecting
- female
-
pre-hatching/birth
-
provisioning
- female
-
protecting
- female
-
provisioning
-
pre-weaning/fledging
-
provisioning
- female
-
provisioning
Lifespan/Longevity
The lifespan of platacanthomyids has not been reported.
Behavior
Very little information is available on the behavior of these rodents. It is known
that
Platacanthomys
is arboreal, and it is assumed that
Typhlomys
is as well.
Platacanthomys
individuals use their large, tufted tails as a balancing organ as they hop between
branches. They build nests of moss and leaves in tree cavities, among branches, or
in rock clefts. One captive
Platacanthomys lasiurus
was lethargic during the day, allowing itself to be handled without struggle, suggesting
nocturnal habits.
Communication and Perception
It is unknown how these rodents communicate. They do have the ability to perceive their world through visual, auditory, tactile, and chemical means, though it is not known how well-developed any of these senses are.
Food Habits
Platacanthomyids reportedly eat leaves, stems, fruit, seeds, and roots.
Predation
There are no reports of predation on platacanthomyids, although it is likely that
small to medium-sized predators, such as large
snakes
,
raptors
, and
mammalian carnivores
, will target these species. Native people of southern China and northern Vietnam
claim that cats will not eat
Typhlomys
species.
Ecosystem Roles
Platacanthomyids are herbivores, meaning that they are at least primary consumers in their ecosystem.
Economic Importance for Humans: Positive
There are no known positive impacts of platacanthomyids on humans, except in their roles in the healthy ecosystems they inhabit.
Economic Importance for Humans: Negative
In parts of India,
Platacanthomys lasiurus
are abundant and are referred to as "pepper rats", because they raid and destroy
pepper crops.
- Negative Impacts
- crop pest
Conservation Status
One of the three species in this family,
Typhlomys chapensis
, the Chapa pygmy dormouse, is listed as critically endangered by the IUCN. This
species is known only from a single locality and therefore is extremely vulnerable
to habitat destruction.
Other Comments
Neither of the current genera in this family are known from the fossil record. The
family is represented by fossils of the extinct genus
Neocometes
, from the early Miocene in Europe.
Additional Links
Contributors
Allison Poor (author), University of Michigan-Ann Arbor, Tanya Dewey (editor), Animal Diversity Web.
- Palearctic
-
living in the northern part of the Old World. In otherwords, Europe and Asia and northern Africa.

- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- oriental
-
found in the oriental region of the world. In other words, India and southeast Asia.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- tropical
-
the region of the earth that surrounds the equator, from 23.5 degrees north to 23.5 degrees south.
- terrestrial
-
Living on the ground.
- forest
-
forest biomes are dominated by trees, otherwise forest biomes can vary widely in amount of precipitation and seasonality.
- endothermic
-
animals that use metabolically generated heat to regulate body temperature independently of ambient temperature. Endothermy is a synapomorphy of the Mammalia, although it may have arisen in a (now extinct) synapsid ancestor; the fossil record does not distinguish these possibilities. Convergent in birds.
- bilateral symmetry
-
having body symmetry such that the animal can be divided in one plane into two mirror-image halves. Animals with bilateral symmetry have dorsal and ventral sides, as well as anterior and posterior ends. Synapomorphy of the Bilateria.
- sexual
-
reproduction that includes combining the genetic contribution of two individuals, a male and a female
- fertilization
-
union of egg and spermatozoan
- internal fertilization
-
fertilization takes place within the female's body
- viviparous
-
reproduction in which fertilization and development take place within the female body and the developing embryo derives nourishment from the female.
- arboreal
-
Referring to an animal that lives in trees; tree-climbing.
- nocturnal
-
active during the night
- motile
-
having the capacity to move from one place to another.
- visual
-
uses sight to communicate
- tactile
-
uses touch to communicate
- acoustic
-
uses sound to communicate
- chemical
-
uses smells or other chemicals to communicate
- herbivore
-
An animal that eats mainly plants or parts of plants.
- folivore
-
an animal that mainly eats leaves.
- frugivore
-
an animal that mainly eats fruit
- granivore
-
an animal that mainly eats seeds
References
Alston, E. 1876. On the classification of the order Glires. Proceedings of the Zoological Society of London : 61-98.
Carleton, M., G. Musser. 1984. Muroid rodents. Pp. 289-379 in Orders and Families of Recent Mammals of the World . New York: John Wiley and Sons.
Chaline, J., P. Mein, F. Petter. 1977. Les grandes lignes d'une classification évolutive des Muroidea. Mammalia , 41: 245-252.
Corbert, G. 1984. Other old world rats and mice. Pp. 666-671 in The Encyclopedia of Mammals . New York: Facts on File.
Ellerman, J. 1940. The Families and Genera of Living Rodents, vol. I . London: British Museum (Natural History).
Helin, S., N. Ohtaishi, L. Houji. 1999. The Mammalian of China . Beijing: China Forestry Publishing House.
IUCN, 2004. "IUCN Red List of Threatened Species" (On-line). Accessed March 25, 2005 at www.redlist.org .
Jansa, S., M. Weksler. 2004. Phylogeny of muroid rodents: relationships within and among major lineages as determined by IRBP gene sequences. Molecular Phylogenetics and Evolution , 31: 256-276.
Michaux, J., A. Reyes, F. Catzeflis. 2001. Evolutionary history of the most speciose mammals: molecular phylogeny of Muroid rodents. Molecular Biology and Evolution , 18(11): 2017-2031.
Miller, G., J. Gidley. 1918. Synopsis of supergeneric groups of rodents. Journal of the Washington Academy of Science , 8: 431-448.
Mudappa, D., A. Kumar, R. Chellam. 2001. Abundance and habitat selection of the Malabar spiny dormouse in the rainforests of the southern Western Ghats, India. Current Science , 80(3): 424-427.
Musser, G., M. Carleton. 2005. Superfamily Muroidea. Mammal Species of the World . Washington: Smithsonian Institution Press.
Norris, R., K. Zhou, C. Zhou, G. Yang. 2004. The phylogenetic position of the zokors (Myospalacinae) and comments on the families of muroids (Rodentia). Molecular Phylogenetics and Evolution , 31: 972-978.
Nowak, R. 1999. Walker's Mammals of the World, v. 2 . Baltimore and London: The Johns Hopkins University Press.
Simpson, G. 1945. The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History , 85: 1-350.
Steppan, S., R. Adkins, J. Anderson. 2004. Phylogeny and divergence-date estimates of rapid radiations in Muroid rodents based on multiple nuclear genes. Systematic Biology , 53(4): 533-553.