Diversity
Scarids, which are widely known as parrotfishes because of their beak-like jaws, include
approximately nine genera and 83 species. They are abundant in tropical reefs around
the world and well known to divers for their striking coloration and noisy feeding
as they crunch on dead coral. Parrotfishes exhibit several types of complex mating
systems that vary more by geographic location than by species (see Reproduction).
They also have considerable ecological impacts on coral reefs through herbivory and
bioerosion (see Ecosystem Roles).
Geographic Range
Parrotfishes are found primarily in tropical waters throughout the Atlantic, Indian,
and Pacific oceans. However, some species inhabit subtropical waters, and some, such
as
Scarus ghobban
, may venture far from reef environments.
- Biogeographic Regions
- nearctic
- palearctic
- oriental
- ethiopian
- neotropical
- australian
- indian ocean
- atlantic ocean
- pacific ocean
- mediterranean sea
Habitat
Most parrotfishes exclusively inhabit offshore coral reefs in tropical regions. However,
a few species feed primarily on sea grasses and are most common in the Caribbean.
Two other species,
Nicholsina denticulate
and
Sparisoma cretensis
, are common over rocky reefs of the Gulf of California and Mediterranean Sea, respectively.
- Habitat Regions
- saltwater or marine
Systematic and Taxonomic History
Scarids are one of several recently derived families (
Acanthuridae
,
Monacanthidae
,
Pomacentridae
,
Blenniidae
,
Siganidae
) capable of exploiting reef algae and small colonial invertebrates. Reef herbivory
was primarily restricted to post-Cretaceous perciformes until these families underwent
rapid evolution during the early Tertiary, 50 to 30 million years ago. The modified
pharyngeal apparatus
, which parrotfishes use to grind dead corals into a digestible paste, is an important
adaptation that has allowed parrotfishes to exploit reef algae and small colonial
invertebrates (see Physical Description and Food Habits for further information).
Brucae and
Randall (1983 and 1985) (in Nelson 1994) created two subfamilies of parrotfishes:
Scarinae
with four genera containing approximately 67 species, and
Sparisomatinae
with five genera and 16 species.
Randall (1983 and 1985) (in Nelson 1994) created two subfamilies of parrotfishes:
Scarinae
with four genera containing approximately 67 species, and
Sparisomatinae
with five genera and 16 species.
{kind=link}
Physical Description
Parrotfishes are characterized by their distinctive beak-like jaws, in which the teeth
are fused together in most species, and a
pharyngeal apparatus
, which acts as a second set of jaws in the throat. In the pharyngeal apparatus,
the teeth are arranged in rows and are highly specialized to grind, crop, and crush
food as it is processed. Parrotfishes have large,
cycloid scales
, usually with 22-24 scales along the lateral line. The dorsal fin has nine spines
and ten soft rays. The anal fin has three spines and nine soft rays, and the pelvic
fins one spine and five soft rays. (Click here to see a
fish diagram
).
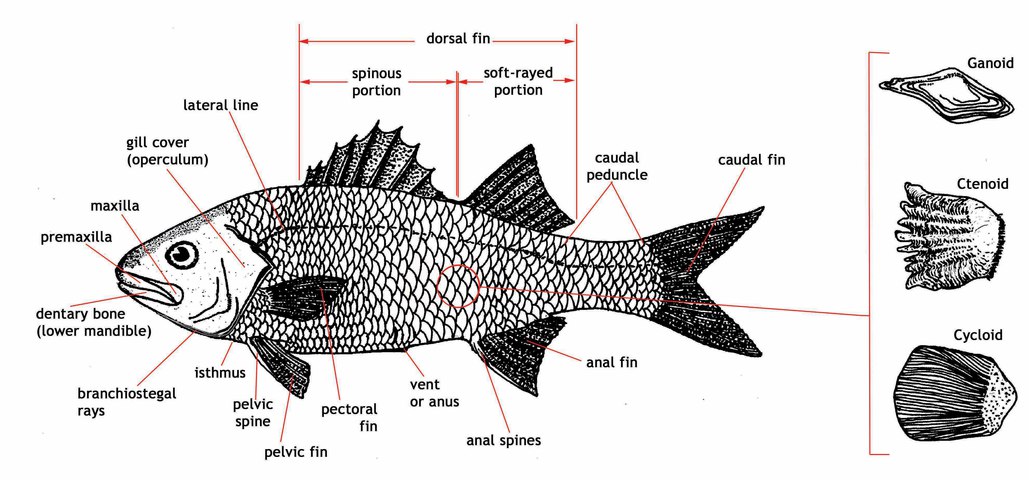{kind=link}
Some parrotfishes have a complex socio-sexual (socially influenced sexual change)
system punctuated by three phases, and each phase change results in a different color
pattern (See Reproduction: Mating Systems for a description of “phases” in parrotfishes).
For instance, juveniles tend to have a drab mixture of browns, grays and blacks,
but as they mature a distinct coloration emerges with the addition of red tones. A
third set of colors is donned by males and by females that have recently undergone
sex change into males. As these males mature, they exhibit bright, intricate patterns
of reds, greens, and blues. This type of color change has been documented in
Scarus
,
Sparisoma
,
Nicholsina
,
Bolbometapon
, and
Cryptotomus
, but there are some monochromic (fishes that do not exhibit sexual color change)
species that exhibit different types of sexual dimorphism.
Scarus coelestinus
and
Scarus coeruleus
in the eastern Pacific and
Scarus niger
in the Indo-West Pacific exhibit no color differences. However, mature males of
Scarus coelestinus
and
Scarus coeruleus
develop more squared-off and prominent foreheads than smaller fish, while
Scarus niger
exhibits no physical differences other than size. Finally, fleshy tips on the upper
and lower lobes of the caudal fin can be observed in mature males of
Scarus rubroviolaceus
, but are poorly developed on small males and females.
- Other Physical Features
- ectothermic
- bilateral symmetry
- Sexual Dimorphism
- male larger
- sexes colored or patterned differently
- male more colorful
- sexes shaped differently
- ornamentation
Development
Tidal currents disperse parrotfish eggs, which begin to hatch approximately 25 hours
after fertilization. Newly hatched larvae begin to feed after three days but the length
of the planktonic stage is unknown. Most parrotfish species develop rapidly and reach
maturity between two and four years.
Reproduction
Parrotfishes utilize some of the most complex and unusual reproduction systems known
to fishes. Males can be either primary, i.e. born male, or secondary, i.e. females
that have undergone sex change. In some species there are no secondary males while
in others all individuals are born female (monandric) and change sex when necessary.
In the most complex systems, species are diandric – both primary and secondary males
exist in the population. In these species, individuals proceed through three distinct
phases, marked by color differences. In fact, the color differences are so pronounced
that for over 200 years researchers regarded some phases as distinct species. Sexually
immature and drab colored juveniles represent the first phase. The second, known as
the initial, phase (IP) can include sexually mature males or females, which are impossible
to tell apart without internal examination or observation during spawning. The terminal
phase (TP) includes only mature males, which display brilliant colors. TP males usually
dominate reproductive activity through a harem-based social system. The death of a
TP male serves as a social cue for an IP female to change sex and behavior. The morphology
and behavior of IP males may also change in response to the death of a TP male. In
some cases IP males attempt to infiltrate a TP male’s harem by masquerading as a female.
In the so called “sneak spawning” attempt IP males follow spawning pairs into the
water column and release a large cloud of gametes at peak spawning in an attempt to
overwhelm fertilization by the TP male. IP males are well equipped to perform “sneak
spawning” as they have larger testes and so are able to produce more gametes, while
TP males have smaller testes and rely on aggression to deter other males.
The type of reproductive behavior described above and whether it involves paired,
foraging group or mass spawning depends on a complex set of behavioral and geographic
factors. For instance, some species, such as
Scarus croicensis
, exhibit a wide range of reproductive behaviors depending on the area in which they
are found. In Panama,
Scarus croicensis
employs a system involving three classes of individuals: territorials, stationeries
and foragers. Territorials are organized into groups that consist of a dominant female,
several subordinate females and usually, but not always, a terminal (TP) male. Paired
spawning occurs within the territory, which both males and females defend. Stationaries
consistently use the same area for spawning but do not defend it, and foragers include
groups of up to 500 individuals, mostly females. In Puerto Rico, initial phase (IP)
and terminal phase (TP) individuals migrate to temporary spawning areas in deep water,
usually in pairs. Finally, in Jamaica
Scarus croicensis
emphasizes aspects of the foraging group system and spawning only takes place in
groups. The three previous examples illustrate the flexibility of the socio-sexual
mating systems found in parrotfishes. The reasons that different aspects of the basic
spawning system manifest in different areas range from population density to competition
for spawning sites and other resources to geographic factors like seasons and water
temperature.
- Mating System
- polygynous
- polygynandrous (promiscuous)
In general, parrotfishes spawn year-round, usually at dusk. However, peak spawning
occurs in summer for many species and there is evidence that some species have defined
non-spawning periods. As discussed above, many species migrate to the outer edges
of the reef to spawn but some spawn within defined territories. There is evidence
that some scarids respond to the lunar cycle during spawning, but in others, spawning
correlates closely with high tide, regardless of the time of the lunar month. In species
that spawn several times during the day, the tidal cycle is followed closely since
this is the optimal time for egg dispersal.
- Key Reproductive Features
- iteroparous
- seasonal breeding
- year-round breeding
- sequential hermaphrodite
- sexual
- fertilization
- oviparous
There is no evidence of parental behavior in parrotfishes.
- Parental Investment
- no parental involvement
Lifespan/Longevity
The maximum age of most parrotfishes is less than 20 years and most live less than
five years. There is a general trend in the scarids for larger species to live longer.
Subsequently, the largest scarid,
Bolbometopon muricatum
, is the one exception to the 20 year maximum age.
Behavior
Parrotfishes are most well known for their complex social structures. Most are organized
into male-dominated harems but others breed cooperatively or in pairs (see Reproduction:
Mating Systems & General Behavior for more on social systems). Some parrotfishes are
highly territorial while others are mainly nomadic, with the home range increasing
as the size of the fish increases. Large foraging groups of up to 500 individuals
form for spawning and to deter predators while feeding. Parrotfishes feed continuously
throughout the day and seek shelter in reefs at night.
- Key Behaviors
- natatorial
- diurnal
- motile
- territorial
- social
- colonial
- dominance hierarchies
Communication and Perception
Most known forms of communication in parrotfishes are related to reproduction and
are discussed in Reproduction: Mating Systems. However, in some species male coloration
intensifies when defending its territory, which suggests that visual cues are used
to deter invaders.
Food Habits
Parrotfishes are primarily herbivorous, grazing intensively on dead, algae-coated
coral, vegetable material, and in some species sea grasses.
Bump-headed parrotfishes
, which consume significant amounts of live coral, are one exception. Key to the success
of parrotfishes is their ability to take up plant material, detritus and calcareous
sediment and process it through the action of the
pharyngeal jaw
. This chewing mechanism grinds ingested material into a fine paste and breaks down
algal cells, releasing the cellular material for digestion. Like
acanthurids
, parrotfishes form large feeding groups, sometimes with multiple species, to overwhelm
territorial fishes and deter predators.
- Primary Diet
- herbivore
Predation
A unique feature of some parrotfishes is the production of a
mucous envelope
at night before resting. The envelope takes about 30 minutes to construct and is
open at both ends to allow water flow. The secreted envelope is foul smelling and
tasting, which may serve to deter nighttime predators that hunt by scent. Most parrotfishes
seek out caves and ledges in the reef for protection at night, but parrotfishes in
the genus
Cryptotomus
bury themselves in the sand like
wrasses
. After creating a hole in the sand
Cryptotomus
then produces its
mucous nightgown
.
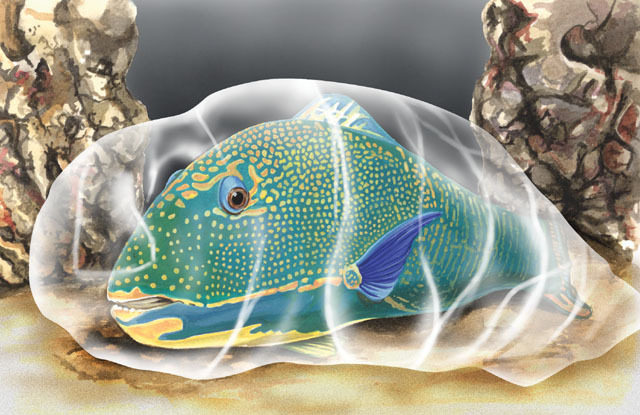{kind=link}
Ecosystem Roles
Parrotfishes have a major impact on coral reefs through intensive grazing and associated
bioerosion. The grazing patterns of large schools of parrotfish have the effect of
selecting for certain species of corals and algae, and preventing algae from choking
out corals. Many parrotfishes feed on calcareous algae (algae that are high in mineral
calcium) growing on dead, exposed coral by biting off chunks and turning them into
a fine paste. This type of grazing contributes significantly to the process of bioerosion
and the creation of sediment on reefs. For instance, it has been calculated that a
single large parrotfish,
Bolbometapon muricatum
(bump-head parrotfish), consumes approximately one cubic meter of coral skeletons
per year, and turns it into fine sediment. In this way large schools of
bump-head parrotfish
determine the fine-scale topography of coral reefs.
A separate ecological consequence of intense herbivory in parrotfishes is the conversion
of plant material into fish flesh. The success of parrotfishes in consuming plant
material unavailable to most other fishes and the large size of parrotfish populations
makes them an important part of the predatory food chain.
- Ecosystem Impact
- biodegradation
Economic Importance for Humans: Positive
In the Bahamas, the scales of some parrotfishes are used for decorating basketwork
and shellflower arrangements, but the fish are not consumed. In other areas, parrotfishes
are sometimes taken as food, but their flesh can be dangerous to humans as a result
of accumulated ciguatera toxins.
- Positive Impacts
- food
- body parts are source of valuable material
- research and education
Economic Importance for Humans: Negative
Several species, such as
blue parrotfish
and one Indo-Pacific species, have caused ciguatera (fish poisoning sickness) in
humans, which can be fatal.
- Negative Impacts
- injures humans
Conservation Status
One scarid,
Scarus guacamaia
(rainbow parrotfish), is listed as vulnerable to extinction.
Other Comments
The fossil history of scarids dates back to the lower Tertiary and Eocene epochs.
Additional Links
Contributors
R. Jamil Jonna (author), Animal Diversity Web.
- Nearctic
-
living in the Nearctic biogeographic province, the northern part of the New World. This includes Greenland, the Canadian Arctic islands, and all of the North American as far south as the highlands of central Mexico.

- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Palearctic
-
living in the northern part of the Old World. In otherwords, Europe and Asia and northern Africa.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- oriental
-
found in the oriental region of the world. In other words, India and southeast Asia.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Ethiopian
-
living in sub-Saharan Africa (south of 30 degrees north) and Madagascar.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Neotropical
-
living in the southern part of the New World. In other words, Central and South America.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Australian
-
Living in Australia, New Zealand, Tasmania, New Guinea and associated islands.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Atlantic Ocean
-
the body of water between Africa, Europe, the southern ocean (above 60 degrees south latitude), and the western hemisphere. It is the second largest ocean in the world after the Pacific Ocean.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Pacific Ocean
-
body of water between the southern ocean (above 60 degrees south latitude), Australia, Asia, and the western hemisphere. This is the world's largest ocean, covering about 28% of the world's surface.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- saltwater or marine
-
mainly lives in oceans, seas, or other bodies of salt water.
- pelagic
-
An aquatic biome consisting of the open ocean, far from land, does not include sea bottom (benthic zone).
- reef
-
structure produced by the calcium carbonate skeletons of coral polyps (Class Anthozoa). Coral reefs are found in warm, shallow oceans with low nutrient availability. They form the basis for rich communities of other invertebrates, plants, fish, and protists. The polyps live only on the reef surface. Because they depend on symbiotic photosynthetic algae, zooxanthellae, they cannot live where light does not penetrate.
- coastal
-
the nearshore aquatic habitats near a coast, or shoreline.
- ectothermic
-
animals which must use heat acquired from the environment and behavioral adaptations to regulate body temperature
- bilateral symmetry
-
having body symmetry such that the animal can be divided in one plane into two mirror-image halves. Animals with bilateral symmetry have dorsal and ventral sides, as well as anterior and posterior ends. Synapomorphy of the Bilateria.
- sexual ornamentation
-
one of the sexes (usually males) has special physical structures used in courting the other sex or fighting the same sex. For example: antlers, elongated tails, special spurs.
- polygynous
-
having more than one female as a mate at one time
- polygynandrous
-
the kind of polygamy in which a female pairs with several males, each of which also pairs with several different females.
- iteroparous
-
offspring are produced in more than one group (litters, clutches, etc.) and across multiple seasons (or other periods hospitable to reproduction). Iteroparous animals must, by definition, survive over multiple seasons (or periodic condition changes).
- seasonal breeding
-
breeding is confined to a particular season
- year-round breeding
-
breeding takes place throughout the year
- protogynous
-
condition of hermaphroditic animals (and plants) in which the female organs and their products appear before the male organs and their products
- sexual
-
reproduction that includes combining the genetic contribution of two individuals, a male and a female
- fertilization
-
union of egg and spermatozoan
- external fertilization
-
fertilization takes place outside the female's body
- oviparous
-
reproduction in which eggs are released by the female; development of offspring occurs outside the mother's body.
- natatorial
-
specialized for swimming
- diurnal
-
- active during the day, 2. lasting for one day.
- motile
-
having the capacity to move from one place to another.
- territorial
-
defends an area within the home range, occupied by a single animals or group of animals of the same species and held through overt defense, display, or advertisement
- social
-
associates with others of its species; forms social groups.
- colonial
-
used loosely to describe any group of organisms living together or in close proximity to each other - for example nesting shorebirds that live in large colonies. More specifically refers to a group of organisms in which members act as specialized subunits (a continuous, modular society) - as in clonal organisms.
- dominance hierarchies
-
ranking system or pecking order among members of a long-term social group, where dominance status affects access to resources or mates
- visual
-
uses sight to communicate
- tactile
-
uses touch to communicate
- visual
-
uses sight to communicate
- tactile
-
uses touch to communicate
- chemical
-
uses smells or other chemicals to communicate
- biodegradation
-
helps break down and decompose dead plants and/or animals
- food
-
A substance that provides both nutrients and energy to a living thing.
- herbivore
-
An animal that eats mainly plants or parts of plants.
References
Berg, L. 1958. System Der Rezenten und Fossilen Fischartigen und Fische . Berlin: VEB Deutscher Verlag der Wissenschaften.
Böhlke, J., C. Chaplin. 1994. Fishes of the Bahamas and Adjacent Tropical Waters . Wynnewood, Pa: Published for the Academy of Natural Sciences of Philadelphia by Livingston.
Choat, H., D. Bellwood. 1998. Wrasses & Parrotfishes. Pp. 209-210 in Encyclopedia of Fishes – second edition . San Diego, CA: Academic Press.
Choat, J., D. Robertson. 2002. Age-Based Studies. Pp. 63-67 in Coral Reef Fishes: Dynamics and Diversity in a Complex Ecosystem . San Diego, CA: Academic Press.
Harmelin-Vivien, M. 2002. Energetics and Fish Diversity on Coral Reefs. Pp. 268-269 in Coral Reef Fishes: Dynamics and Diversity in a Complex Ecosystem . San Diego, CA: Academic Press.
Nelson, J. 1994. Fishes of the World – third edition . New York, NY: John Wiley and Sons.
The World Conservation Union, 2002. "IUCN 2002" (On-line). IUCN Red List of Threatened Species. Accessed August 02, 2003 at http://www.iucnredlist.org/ .
Thresher, R. 1984. Reproduction in Reef Fishes . Neptune City, NJ: T.F.H. Publications.
Wheeler, A. 1985. The World Encyclopedia of Fishes - second edition . London: Macdonald.