Diversity
Wrasses (the family Labridae), are the most abundant and conspicuous fishes on tropical
reefs around the world. Wrasses also comprise an important element of the coldwater
fish population on temperate reefs. They are second largest family of marine fishes
and the third largest family in the
Perciformes
order, containing approximately 60 genera and roughly 500 species. Wrasses appear
in a diverse range of colors, shapes, and sizes, often varying considerably within
individual species (see Physical Description). This morphological diversity is matched
by the wide variety of prey consumed. Wrasses fill the roles of piscivores, zooplanktivores,
molluscivores, herbivores, planktivores, polychaete predators, decapod crab predators,
and coral predators, as well as many others (see Food Habits). Many wrasses are organized
into harem-based social systems and hermaphroditism is common (see Reproduction: Mating
Systems). Finally, as suggested by their diverse food habits, wrasses fill many important
ecological roles on reefs of tropical and temperate regions around the world.
Geographic Range
Wrasses occupy all tropical seas and penetrate considerable distances into temperate
waters, reaching as far north as Norway. Many temperate species in the genera
Oxyjulius
,
Tautoga
,
Tautogolabrus
,
Semicossyphus
, and
Labrus
can be found in both the Atlantic and Pacific Oceans. Wrasses are most highly concentrated
off the coasts of Australia where about 165 species and 42 genera are represented.
- Biogeographic Regions
- nearctic
- palearctic
- oriental
- ethiopian
- neotropical
- australian
- oceanic islands
- indian ocean
- atlantic ocean
- pacific ocean
- mediterranean sea
Habitat
Wrasses can be found in a wide variety of habitats, such as tidal pools, grass beds,
rocky or coral reefs, or open sand bottoms. Many wrasses prefer specific environments.
Doratonotus
, for example, prefer turtle grass beds,
Hemipteronotus
, mixed turtle grass and sandy patch areas, and
hogfishes
, weed-covered rocky flats. Plankton feeders, such as
Clepticus
, often concentrate in large schools at reef fronts, reef gaps, or other areas where
plankton is concentrated in the water column. However, some species, such as the
slippery dick
, can be found in a broad range of habitats.
- Habitat Regions
- temperate
- tropical
- saltwater or marine
- Aquatic Biomes
- benthic
- reef
- coastal
- brackish water
- Other Habitat Features
- estuarine
- intertidal or littoral
Systematic and Taxonomic History
Since male, female, and juvenile wrasses come in radically different colors and shapes
(see Physical Description) and are conspicuous in shallow waters around the world,
investigators initially named far too many distinct wrasse species. The number of
species has been steadily declining but wrasses are still the third largest family
of
perciform
fishes and the second largest family of marine fishes. Although many subfamilies
and tribes are recognized, only one is monophyletic: tribe
Cheilinini
with 21 species (Westneat 1993 from Nelson 1994).
Physical Description
Most wrasses are quite small, usually below 20 cm. The smallest species,
Minilabrus striatus
of the Red Sea, reaches a maximum length of only 4.5 cm. The genera
Pseudocheilinus
and
Doratonotus
contain several other dwarf wrasses. One species,
Conniella apterygia
, is so small that it lacks even pelvic fins and a supporting skeleton. The largest
wrasse,
Cheilinus undulatus
, can reach a length of about 2.3 m and weighs more than 150 kg. Wrasses are most
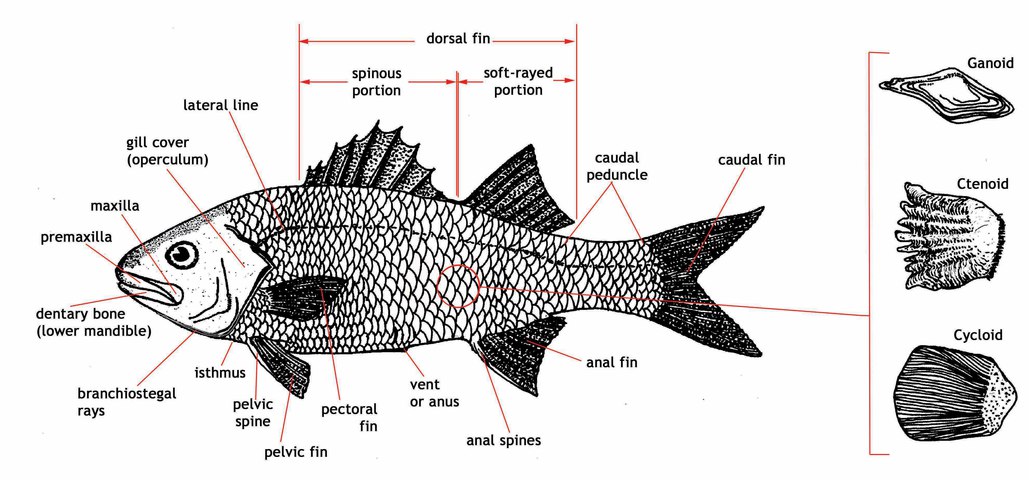
easily identified by their pointed snouts and prominent canine teeth in the front
of the jaws, which often project forward. Wrasses characteristically have a protractile
mouth,
cycloid scales
, and a single continuous dorsal fin lacking an obvious notch between the soft and
spiny portions. The lateral line may be continuous or interrupted. (Click here to
see a
fish diagram
).
{kind=link}
Wrasses display myriad colors and shapes.
Razorfishes
are elongate and laterally compressed, while members of
Cheilinus
,
Choerodon
, and many of
Bodianus
are large and stocky. However, most are elongate and tapered at both ends, often
referred to as “cigar-shaped.” Cigar-shaped fishes are found in the genera
Thalassoma
,
Halichoeres
, and
Labroides
. Often, there is considerable diversity of colors and shapes within individual species.
As in
parrotfishes
, some wrasses progress through “phases” (see Reproduction: Mating Systems), and each
phase corresponds with a change in morphology (shape and color). Dominant males (and
sometimes females) are the most distinctly colored, with complex patterns of red,
yellow, green, blue and black. Subordinate males and females are smaller than dominant
individuals and are often drab-colored with cryptic patterns. Juveniles range in coloration
from bright yellow and orange to drab gray and brown, and some have camouflaging patterns.
(See Reproduction: Mating Systems for details). Some wrasses exhibit
sexual dimorphism
.
{kind=link}
- Other Physical Features
- ectothermic
- bilateral symmetry
- polymorphic
- Sexual Dimorphism
- male larger
- sexes colored or patterned differently
- male more colorful
- sexes shaped differently
Development
Wrasses occupy a wide range of water temperatures and incubation time is directly
affected by water temperature. In laboratory experiments incubation took approximately
24 hours at 27˚C. The planktonic stage is estimated to be around one month, although
very little is known about this stage. The age or size at which individuals reach
sexual maturity depends on the maximum size of the species.
Reproduction
Like
parrotfishes
, many wrasses utilize some of the most complex and unusual reproduction systems known
to fishes. Males can be either primary (born male), or secondary (females that have
undergone sex change). In some species there are no secondary males while in others
all individuals are born female (monandric) and change sex when necessary. In the
most complex systems, species are diandric – both primary and secondary males exist
in the population. In these species, individuals proceed through three distinct phases,
marked by color differences. In fact, the color differences are so pronounced that
for over 200 years researchers regarded some phases as distinct species. Sexually
immature juveniles represent the first phase. The second, known as the initial, phase
(IP) can include sexually mature males or females, which are impossible to tell apart
without internal examination or observation during spawning. IP males and females
may group spawn in some species. The terminal phase (TP) includes only mature males,
which display brilliant colors. TP males usually dominate reproductive activity through
a harem-based social system. The death of a TP male serves as a social cue for an
IP female to change sex and behavior. The morphology of IP males may also change in
response to the death of a TP male. In some cases, IP males attempt to fertilize IP
females by following a TP male and IP female pair during spawning. In this behavior,
called “streaking,” IP males follow the pairs at peak spawning and release a large
cloud of gametes in an attempt to overwhelm fertilization by the TP male. This is
thought to increase the fecundity (ability to produce offspring) of IP males. IP males
are well equipped to perform streaking as they have larger gonads and so are able
to produce more gametes, while TP males have smaller testes and rely on aggression
to deter other males. The larger volume of milt (gametes) produced by IP males is
related to group spawning events with IP females, in which competition for fertilization
is intense and more milt is needed.
Some specific examples of wrasse mating systems demonstrate the complexity and variation
of the phase system described above. For instance, the
cleaner wrasse
, which is monandric (all individuals are born female), forms harems that are held
together by male aggression towards subordinate females. With the death of the dominant
male, subordinate females jockey for position and the newly dominant female adopts
aggressive male behavior within a few hours. Each individual moves a step up in the
dominance hierarchy and the last position is filled by a juvenile. If the newly dominant
female is able to withstand attempts by neighboring males to take over the vacant
harem, she will become a fully functional male within a two to four days. Some other
harem-forming species are
Cirrhilabrus temminckii
,
Cirrhilabrus jordani
,
Labroides bicolor
,
Hemipteronotus splendens
,
Pseudocheilinus hexataenia
and
Macropharyngodon moyeri
. The Caribbean species
Halichoeres garnoti
is also monandric, but individuals do not exhibit territoriality or conspicuous dominance
relationships, nor do they use aggressive actions to maintain sexual state. Instead,
size or some size-related factor determines which individual will fill the male role.
In
Halichoeres garnoti
males are larger than females and both sexes behave similarly. While these examples
focus on the mating extremes of wrasses, most species fall between the systems of
the
cleaner wrasse
and
Halichoeres garnoti
in terms of the influence of social control on sex reversal. Other hermaphroditic
but non-harem-forming species include
Halichoeres bivittatus
and
Halichoeres poeyi
,
Halichoeres maculipinna
and possibly
Thalassoma lunare
. Finally, some species, such as
Oxyjulis californica
and
Crenilabrus melops
, do not follow the phase system at all as they are not hermaphroditic, and there
are probably more non-hermaphroditic species yet to be found.
- Mating System
- polygynous
- polygynandrous (promiscuous)
In tropical wrasses spawning occurs year-round but some temperate species seem to
restrict spawning to warmer parts of the year. Spawning typically occurs along the
outer edge of patch reefs or along the outer edges of more extensive reef complexes.
The correlation between spawning and lunar periodicity (the lunar cycle) is sketchy
in some species and non-existent in most that have been investigated. Spawning in
several species corresponds with outgoing tides, however, many species spawn at a
particular time in the day, regardless of tidal patterns. This variation may be due
to local conditions. For instance, in areas where tidal forces are weak, factors like
time of day or light intensity may have more influence. However, evidence from different
species on the same reef suggests that temporal (measured time) differences in spawning
evolved to decrease the probability of hybridization with other species.
Wrasses may spawn in groups or pairs depending on the species or phase of individuals.
Typically, group or aggregate spawning occurs between initial phase (IP) individuals,
which are diandric (containing male and female IP individuals). However, in some species,
such as
Thalassoma cupido
,
Thalassoma lucasanum
, and
Halichoeres bivitattus
, terminal phase (TP) males have been observed participating in group spawning. The
size of the spawning groups ranges from a dozen to several hundred individuals. Males
outnumber females, sometimes by as much as ten to one. Paired spawning is found in
many, if not all, tropical wrasses and involves a TP male and IP female. In rare cases,
IP individuals also spawn in pairs. Most species defend small territories only during
spawning. Currently
Anampses cuvieri
is the only known species of tropical wrasse to produce demersal eggs (eggs laid
on the bottom as opposed to being released in the water column). Demersal spawning
of
Anampses cuvieri
was only observed in captivity and still needs to be confirmed, but work on other
species of this genus seems to support this observation.
- Key Reproductive Features
- iteroparous
- year-round breeding
- gonochoric/gonochoristic/dioecious (sexes separate)
- sequential hermaphrodite
- sexual
- fertilization
- oviparous
Some temperate wrasse species, such as the
ballan wrasse
and
Anampses cuvieri
, are demersal nest builders. The nests are usually made out of plant material and
the male guards the eggs after they are deposited.
- Parental Investment
- male parental care
Lifespan/Longevity
No information was found concerning the lifespan of wrasses but, in general, reef
species live between three and five years.
Behavior
A characteristic feature of wrasses is their form of propulsion, which relies almost
entirely on the pectoral fins. In what is termed “pectoral fins only” propulsion or
labriform locomotion, the fish bounces through the water column using the pectoral
fins and the caudal fin (tail) is only used when a burst of speed is needed. Wrasses
are also strongly diurnal (only active during the daytime) and, like
parrotfishes
, many bury themselves in the sand or seek crevices to hide in at night. Interestingly,
observations of wrasses in captivity seem to suggest a rapid eye movement (REM) stage
while sleeping. REM sleep is usually associated with dreaming in “higher” vertebrates.
Wrasses may forage individually, in pairs, or in large schools depending on the species.
- Key Behaviors
- natatorial
- diurnal
- parasite
- motile
- territorial
- social
- dominance hierarchies
Communication and Perception
Most wrasses rely on vision to find their prey. Visual recognition may also be important
for terminal phase (TP) males to identify harem members. Although TP males are susceptible
to streaking attempts by initial phase (IP) males (see Reproduction: Mating Systems),
no IP males have been found in harem-forming species. This suggests that IP males
are unable to mimic IP females, despite very similar morphology.
- Communication Channels
- visual
- Other Communication Modes
- mimicry
Food Habits
Many wrasses are specialized and voracious feeders, as reflected by the highly variable
skull and body shape, modified
pharyngeal jaw
, and prominent canines. The type of nourishment ranges widely: fish, ectoparasites,
mollusks, polychaete worms, decapod crabs, corals, coral mucous, amphipods, various
echinoderms, plankton, and several types of vegetation. Many small wrasses follow
larger fishes and exploit any benthic (reef bottom) disturbances that help to reveal
the well-camouflaged invertebrates. A considerable number are plankton feeders, forming
schools in reef gaps, reef fronts or other areas with current. The food habits of
cleaner wrasses
are probably most well known.
Cleaner wrasses
remove mucous, parasites and scales from the bodies of larger fishes. Cleaning is
not limited to the
Labroides
genus however; young
bluehead
and young
Spanish hogfish
in the Bahamas have also been observed cleaning larger fishes. Finally, some piscivorous
(fish-eating) wrasses mimic harmless fishes (Randall and Kuiter, 1989 in Nelson, 1994).
{kind=link}
- Primary Diet
-
carnivore
- piscivore
- eats eggs
- eats body fluids
- eats non-insect arthropods
- molluscivore
- herbivore
- omnivore
Predation
Many juvenile wrasses are cryptically colored to avoid predation while others find
protection among the tentacles of
sea anemones
. Nearly all adult wrasses bury themselves in sand at night to avoid predators. A
few species seek out reef crevices and produce a foul-smelling
mucous bag
to deter predators while sleeping. Razorfishes (
Hemipteronotus
,
Xyrichtys
) also use the sand for protection during the day by diving into the bottom. Razorfishes
are apparently quite agile in this environment, sometimes resurfacing several meters
from the point of entry.
{kind=link}
- Anti-predator Adaptations
- mimic
- cryptic
Ecosystem Roles
The ecological role of
cleaner wrasses
of the Indo-Pacific region provides a good example of the complexity of seemingly
mutualistic relationships between fishes. Typically, cleaner fishes are elaborately
colored and perform displays over a patch of reef while larger fish approach and assume
a relaxed posture. Cleaner fishes are commonly thought to benefit the host by removing
dead or damaged tissue and ectoparasites. Accordingly, investigators reported higher
recovery rates for wounded fish in the presence of cleaners. However, in experiments
where all cleaners were removed from an environment there was no incidence of fishes
leaving the area or becoming particularly unhealthy. Further, when levels of parasitic
infections are high the host benefits from cleaning but when infection levels are
low, which they usually are, some cleaners feed on healthy tissue, such as scales,
pieces of fin, mucous, or in some cases the eggs of other reef fishes. Despite these
parasitic qualities of the relationship, fishes being cleaned have a positive response
to the tactile stimulation from cleaners, suggesting that some cleaners are mildly
beneficial while others have taken advantage of the cleaning arrangement.
The relationship between wrasse species and their invertebrate prey is a spectacular
example of coevolution. As invertebrates have developed anti-predator adaptations,
such as spines, toxins, heavy armor, and adherence to the substrate, wrasses have
evolved simultaneously. Some physical changes include the development of strong, hard
beaks and a second set of strong teeth in the throat (
pharyngeal jaw
), making it possible to crush hard-shelled invertebrates. A conspicuous behavioral
adaptation is “following behavior.” As larger fish disturb the substrate, some wrasses
follow close behind to capture exposed invertebrates. Other small wrasses have become
adept at combing the reef for invertebrates too small for most fishes to prey upon.
Finally, some wrasses use their snouts to flip rocks and pieces of coral to expose
hidden invertebrates.
- Ecosystem Impact
- parasite
- some fishes
- many fish species
Economic Importance for Humans: Positive
Wrasses from the
Coris
genera are popular aquarium fishes and two species from the Atlantic coast of North
America, the
cunner
and the
tautog
, are valued as commercial and sport fish. Some other medium to large wrasses are
popular food fishes as well.
Economic Importance for Humans: Negative
No specific information was found concerning any negative impacts to humans.
Conservation Status
Four labrid species are listed as vulnerable:
Cheilinus undulates
,
Lachnolaimus maximus
,
Thalassoma ascensionis
, and
Xyrichtys virens
.
Other Comments
The fossil history of Labridae dates back to the lower Tertiary and Paleocene epochs.
Additional Links
Contributors
R. Jamil Jonna (author), Animal Diversity Web.
- Nearctic
-
living in the Nearctic biogeographic province, the northern part of the New World. This includes Greenland, the Canadian Arctic islands, and all of the North American as far south as the highlands of central Mexico.

- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Palearctic
-
living in the northern part of the Old World. In otherwords, Europe and Asia and northern Africa.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- oriental
-
found in the oriental region of the world. In other words, India and southeast Asia.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Ethiopian
-
living in sub-Saharan Africa (south of 30 degrees north) and Madagascar.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Neotropical
-
living in the southern part of the New World. In other words, Central and South America.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Australian
-
Living in Australia, New Zealand, Tasmania, New Guinea and associated islands.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- oceanic islands
-
islands that are not part of continental shelf areas, they are not, and have never been, connected to a continental land mass, most typically these are volcanic islands.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Atlantic Ocean
-
the body of water between Africa, Europe, the southern ocean (above 60 degrees south latitude), and the western hemisphere. It is the second largest ocean in the world after the Pacific Ocean.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Pacific Ocean
-
body of water between the southern ocean (above 60 degrees south latitude), Australia, Asia, and the western hemisphere. This is the world's largest ocean, covering about 28% of the world's surface.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- temperate
-
that region of the Earth between 23.5 degrees North and 60 degrees North (between the Tropic of Cancer and the Arctic Circle) and between 23.5 degrees South and 60 degrees South (between the Tropic of Capricorn and the Antarctic Circle).
- tropical
-
the region of the earth that surrounds the equator, from 23.5 degrees north to 23.5 degrees south.
- saltwater or marine
-
mainly lives in oceans, seas, or other bodies of salt water.
- benthic
-
Referring to an animal that lives on or near the bottom of a body of water. Also an aquatic biome consisting of the ocean bottom below the pelagic and coastal zones. Bottom habitats in the very deepest oceans (below 9000 m) are sometimes referred to as the abyssal zone. see also oceanic vent.
- reef
-
structure produced by the calcium carbonate skeletons of coral polyps (Class Anthozoa). Coral reefs are found in warm, shallow oceans with low nutrient availability. They form the basis for rich communities of other invertebrates, plants, fish, and protists. The polyps live only on the reef surface. Because they depend on symbiotic photosynthetic algae, zooxanthellae, they cannot live where light does not penetrate.
- coastal
-
the nearshore aquatic habitats near a coast, or shoreline.
- brackish water
-
areas with salty water, usually in coastal marshes and estuaries.
- estuarine
-
an area where a freshwater river meets the ocean and tidal influences result in fluctuations in salinity.
- intertidal or littoral
-
the area of shoreline influenced mainly by the tides, between the highest and lowest reaches of the tide. An aquatic habitat.
- ectothermic
-
animals which must use heat acquired from the environment and behavioral adaptations to regulate body temperature
- bilateral symmetry
-
having body symmetry such that the animal can be divided in one plane into two mirror-image halves. Animals with bilateral symmetry have dorsal and ventral sides, as well as anterior and posterior ends. Synapomorphy of the Bilateria.
- polymorphic
-
"many forms." A species is polymorphic if its individuals can be divided into two or more easily recognized groups, based on structure, color, or other similar characteristics. The term only applies when the distinct groups can be found in the same area; graded or clinal variation throughout the range of a species (e.g. a north-to-south decrease in size) is not polymorphism. Polymorphic characteristics may be inherited because the differences have a genetic basis, or they may be the result of environmental influences. We do not consider sexual differences (i.e. sexual dimorphism), seasonal changes (e.g. change in fur color), or age-related changes to be polymorphic. Polymorphism in a local population can be an adaptation to prevent density-dependent predation, where predators preferentially prey on the most common morph.
- polygynous
-
having more than one female as a mate at one time
- polygynandrous
-
the kind of polygamy in which a female pairs with several males, each of which also pairs with several different females.
- iteroparous
-
offspring are produced in more than one group (litters, clutches, etc.) and across multiple seasons (or other periods hospitable to reproduction). Iteroparous animals must, by definition, survive over multiple seasons (or periodic condition changes).
- year-round breeding
-
breeding takes place throughout the year
- protogynous
-
condition of hermaphroditic animals (and plants) in which the female organs and their products appear before the male organs and their products
- sexual
-
reproduction that includes combining the genetic contribution of two individuals, a male and a female
- fertilization
-
union of egg and spermatozoan
- external fertilization
-
fertilization takes place outside the female's body
- oviparous
-
reproduction in which eggs are released by the female; development of offspring occurs outside the mother's body.
- male parental care
-
parental care is carried out by males
- natatorial
-
specialized for swimming
- diurnal
-
- active during the day, 2. lasting for one day.
- parasite
-
an organism that obtains nutrients from other organisms in a harmful way that doesn't cause immediate death
- motile
-
having the capacity to move from one place to another.
- territorial
-
defends an area within the home range, occupied by a single animals or group of animals of the same species and held through overt defense, display, or advertisement
- social
-
associates with others of its species; forms social groups.
- dominance hierarchies
-
ranking system or pecking order among members of a long-term social group, where dominance status affects access to resources or mates
- visual
-
uses sight to communicate
- mimicry
-
imitates a communication signal or appearance of another kind of organism
- visual
-
uses sight to communicate
- tactile
-
uses touch to communicate
- chemical
-
uses smells or other chemicals to communicate
- cryptic
-
having markings, coloration, shapes, or other features that cause an animal to be camouflaged in its natural environment; being difficult to see or otherwise detect.
- parasite
-
an organism that obtains nutrients from other organisms in a harmful way that doesn't cause immediate death
- pet trade
-
the business of buying and selling animals for people to keep in their homes as pets.
- food
-
A substance that provides both nutrients and energy to a living thing.
- carnivore
-
an animal that mainly eats meat
- piscivore
-
an animal that mainly eats fish
- molluscivore
-
eats mollusks, members of Phylum Mollusca
- herbivore
-
An animal that eats mainly plants or parts of plants.
- omnivore
-
an animal that mainly eats all kinds of things, including plants and animals
References
Allen, G., D. Robertson. 1994. Fishes of the Tropical Eastern Pacific . San Diego, CA: Academic Press.
Berg, L. 1958. System Der Rezenten und Fossilen Fischartigen und Fische . Berlin: VEB Deutscher Verlag der Wissenschaften.
Böhlke, J., C. Chaplin. 1984. Fishes of the Bahamas and Adjacent Tropical Waters . Wynnewood, Pa: Published for the Academy of Natural Sciences of Philadelphia by Livingston.
Choat, H., D. Bellwood. 1998. Wrasses & Parrotfishes. Pp. 209-213 in Encyclopedia of fishes – second edition . San Diego, CA: Academic Press.
Helfman, G., B. Collete, D. Facey. 1997. The Diversity of Fishes . Malden, MA: Blackwell.
Moyle, P., J. Cech. 2000. Fishes: An Introduction to Ichthyology – fourth edition . Upper Saddle River, NJ: Prentice-Hall.
Nelson, J. 1994. Fishes of the World – third edition . New York, NY: John Wiley and Sons.
The World Conservation Union, 2002. "IUCN 2002" (On-line). 2002 IUCN Red List of Threatened Species. Accessed August 22, 2003 at http://www.iucnredlist.org/ .
Thresher, R. 1984. Reproduction in Reef Fishes . Neptune City, NJ: T.F.H. Publications.
Wainwright, P., D. Bellwood. 2002. Ecomorphology of Feeding in Coral Reef Fishes. Pp. 33-55 in Coral Reef Fishes: Dynamics and Diversity in a Complex Ecosystem . San Diego, CA: Academic Press.
Wheeler, A. 1985. The World Encyclopedia of Fishes - second edition . London: Macdonald.