Diversity
The family
Unionidae
is commonly referred to as pearly mussels, naiads, or unionids. Although no full
accounts for the family
Unionidae
exist and the exact number is currently disputed, researchers agree that the order
Unionoida
includes around 1,000 species worldwide (Bauer 2001a). Charles Torrey Simpson described
1,172 species in 1900 and 1,337 in 1914. A more recent account by Fritz Haas (1969)
combined over 4,000 names into just 837 recognized species (Graf & Cummings 2002).
The
Unionidae
are acephalic (no head), bivalved mollusks usually with the beak (the elevated portion
of the dorsal margin) slightly anterior. When present, the pseudocardinal teeth are
generally anterior to the beak. The lateral teeth, generally posterior to the beak,
are parallel to the hinge line. The species in this family have a foot rather than
a byssus, fibrous structures found in other mussel families. Along with
Margaritiferidae
, another family included in the order
Unionoida
,
Unionidae
does not have true siphons (True siphons are formed when tissues between the inhalent
and exhalent openings are fused and mantle aperatures are elongated). Unlike the
family
Margaritiferidae
, the inhalant aperture (opening in the posterior end of the mantle border where water
enters the mussel) of
Unionidae
has unbranched papillae (bumps). Individuals vary in shape, size and coloration.
Adult individuals can range from 30 to 250 mm.
Geographic Range
Members of the family
Unionidae
occur in North America, Europe, Asia, Africa, and the Indonesian Archipelago (Graf
and Cummings, 2002) and can thrive in tropical to temperate climates. The most diversity
is in North America, where there are approximately 286 species (Turgeon et al., 1998),
mainly east of the Rocky Mountains (Jennings, 2000). The nearly 300 species in North
America are grouped into 49 genera, which make up two subfamilies:
Unioninae
and
Ambleminae
(Graf, 2002). The genus
Elliptio
is an example of this broad diversity. Only two species are found in the interior
United States (Mississippi River basin), and the majority of the species (36 currently
recognized) are found in the rivers of the southeastern Atlantic coastal plain (Watters,
2001a). Historical documentation describes mussels paving the beds of the Ohio and
Wabash rivers (Warren, 2000).
- Biogeographic Regions
- nearctic
- palearctic
- oriental
- australian
- Other Geographic Terms
- holarctic
Habitat
Unionids are found in various permanent freshwater sources such as lakes, streams,
and rivers. The family
Unionidae
is not found in high mountain lakes, probably due to a lack of proper fish hosts
for the glochidia or poor nutrient supply (Smith 2001). Most species are generally
found where there are coarse substrates like sand or gravel (Smith 2001) however,
the predictive value of substrate has been questioned (Strayer and Ralley, 1993).
In Michigan, different mussel distributions may more strongly tied to surface geology
in the streams (Strayer, 1983). Constantly shifting substrates or stream basins composed
of solid rock have few mussels. Rivers tend to have a more abundant food supply and
higher dissolved oxygen content than bodies of water with little or no current. Large
rivers tend to contain a wider diversity of mussel species and larger populations
than smaller streams (Cummings & Mayer 1992). Watters (1992) found as the area of
a drainage basin increases, so does the fish diversity. This relationship is likely
due to the increased diversity in habitat for fish. Watters (1992) also found a linear
correlation between fish diversity and mussel diversity, likely due to the increase
number of host fish species available.
Because the shell is primarily composed of calcium carbonate, mussels prefer an aquatic
habitat with an alkaline pH, an abundance of calcium, a bound carbon dioxide content
of more than 15 mg/L, and a potassium level less than 7 mg/L. Some species are able
to tolerate an acidic pH for a short time, but eventually the acid will dissolve the
shell and alter the internal chemistry of the visceral mass. Calcium and carbon dioxide
are important for the development of the shell, and potassium appears to be toxic.
Unionids are most abundant in depths less than 2 m, but will populate waters as deep
as 7 m (Smith 2001). The record depth for a
Unionidae
genus was 31 m. At this depth six
Pyganodon grandis
specimens ranging between 7 and 14 years old and less than 53 mm long were collected
from Lake Michigan (Reigle 1967).
- Habitat Regions
- temperate
- tropical
- freshwater
- Aquatic Biomes
- benthic
- lakes and ponds
- rivers and streams
- Other Habitat Features
- intertidal or littoral
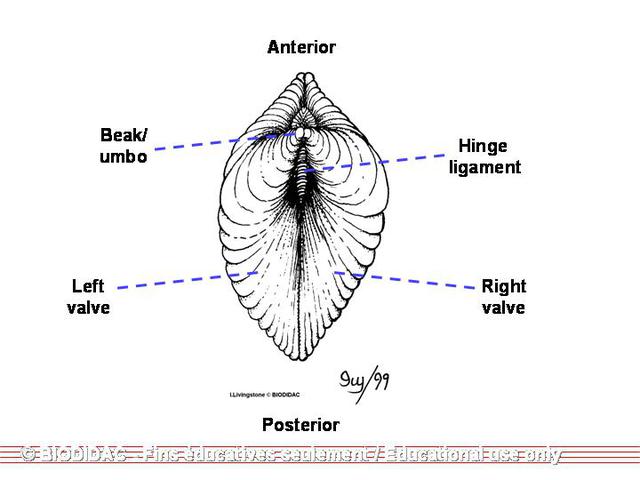
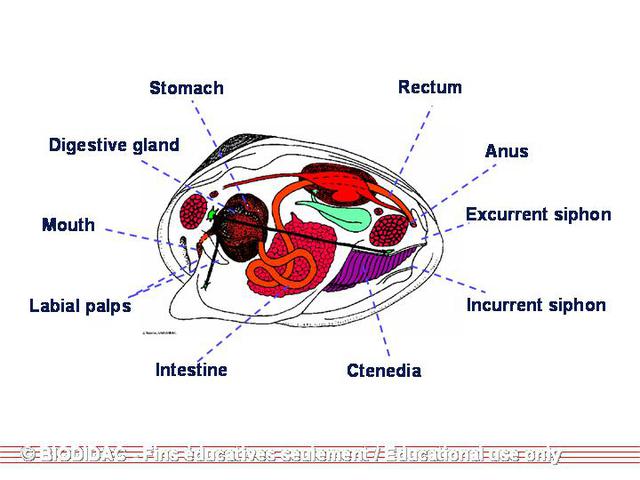
Physical Description
In general, members of the
Unionidae
are acephalic, have two calcium carbonate/organic
shells
called "valves" (bivalved) attached at the hinge by an elastic ligament. They have
an
umbo
(beak) along the dorsal margin and slightly anterior to the hinge and are bilaterally
symmetrical along a plane running between the two valves. Individuals do not have
true siphons. Instead, they have two to three openings in the mantle along the posterior
margin that act as the inhalant and exhalant apertures (Smith 2001). These openings
are either papillated (bumpy) or crenulated (grooved) along the external margin. Under
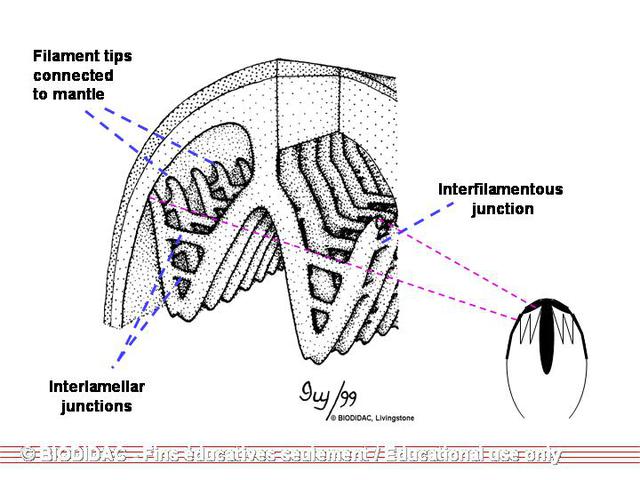
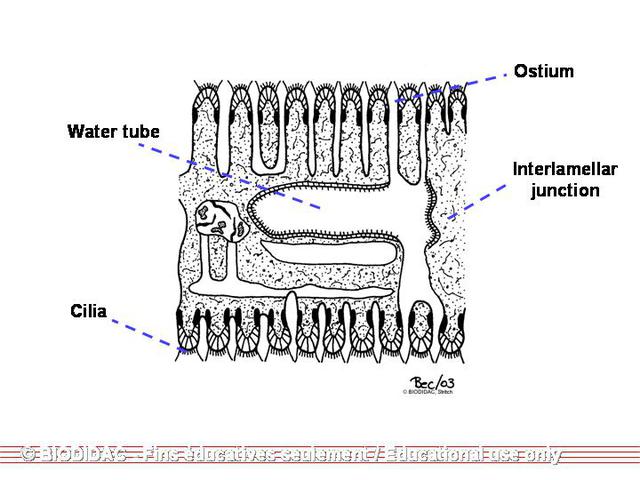
each mantle is a gill made up of two demibranchs. Each demibranch is composed of two
lamellae
fused at the ventral surface but open at the dorsal surface forming a “W.” Each lamella
is lined vertically with compact
water tubes
, which are closed at the bottom but open into a larger, shared cavity at the top
called the suprabranchial chamber. These water tubes are characteristic of
Unionidae
but not
Margaritiferidae
. The ax-shaped
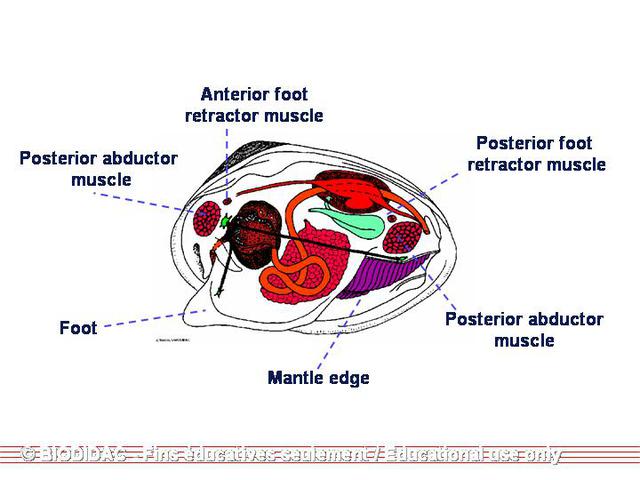
foot
is found on the anterior end of the organism and between the demibranchs in the two
valves. The majority of the median visceral mass in the posterior portion of the organism
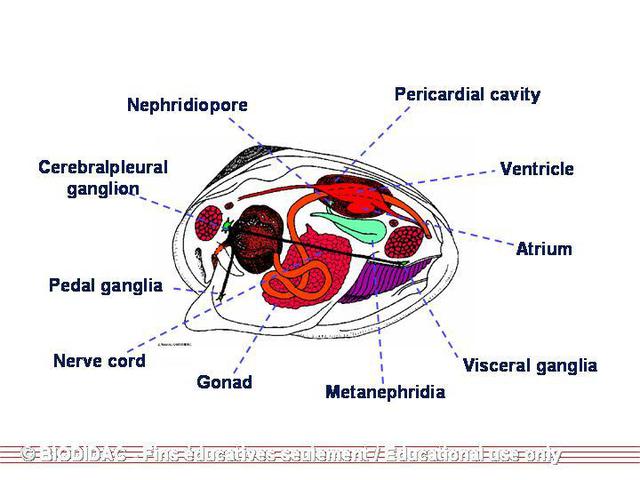
is primarily dorsal and not as confined in the anterior portion (Smith 2001). Unionids
have a simple sensory system. Their
nervous system
is comprised of three pairs of ganglia: cerebropleural, pedal, and visceral. With
one on each side of the esophagus, the cerebropleural ganglia are located on the posterior
side of the anterior adductor muscle and are connected by a short commissure. In the
foot and fused is the pair of pedal ganglia and anterior to the posterior adductor
muscle is the partially fused visceral ganglia. The ganglia are connected by long
commissures and each pair is the source of the nerve fibers for the surrounding organs
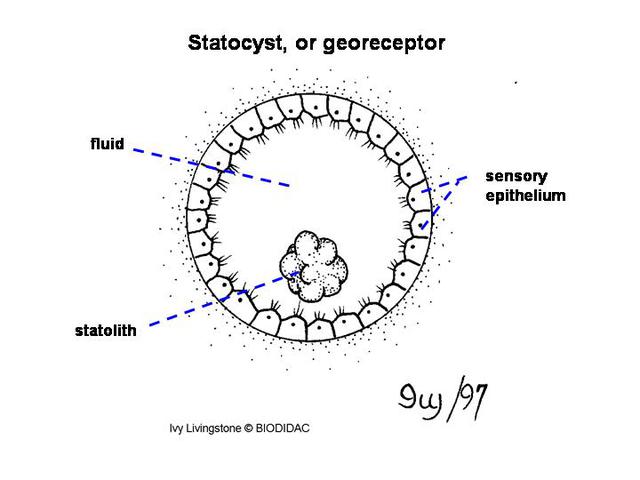
(Smith 2001). Near the pedal ganglia is a pair of
statocysts
, which are ovid or spherical. These statocysts are filled with fluid and lined with
sensory cells. They also contain a solid sphere called a statolith (Smith 2001). These
mussels generally have closed statocysts and a single statolith (Meglitsch & Schram
1991). Osphradia are specialized epithelium concentrated in two small regions on
the roof of the cloacal chamber (the posterior end of the suprabranchial chamber in
the gills where it is fused) (Smith 2001). In some species, there is a spot of pigmentation
near the inhalant aperture that may be photoactive (Smith 2001).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Adult unionids can range anywhere from 30 to 250 mm (Smith 2001) in length, and are
just as variable in shape and color. Among the common shapes are triangular, circular,
rhomboidal, quadrate, trapezoidal, and elliptical (Burch 1975). Shape is a general
description; it cannot be heavily relied upon in the identification of species because
it can vary among individuals of the same species. It is not uncommon to have a more
inflated, rounded form of a species found in large rivers, while the larger, more
compressed form of the same species is found in smaller streams and lakes where currents
are not as strong. Many genera in the subfamily
Ambleminae
, tribe
Lampsilini
exhibit sexual dimorphism. In these species, the males are usually bluntly pointed
or squared along the posterior-ventral margin, while females are broadly truncated.
Periostracum colors vary from yellow or tan to shades of green to dark brown or black.
Some have solid rays, broken rays, wavy rays, rays composed of chevrons, or even a
combination of rays and spots. External shell sculpturing can also vary from one species
to another and can be used to distinguish some taxa. Sculptures can be one of several
combinations of ridges and bumps called "nodules" or "pustules." Not all mussel species
have sculpturing. Nearly the entire
Unioninae
subfamily has smooth surfaces with the exception of ridges formed from the concentric
growth rings. Another exterior sculpturing that is relied upon in identification is
beak sculpture. Beak sculptures range from numerous fine concentric ridges to a few
distinctive bars to double-looped or v-shaped ridges. In some cases, the difference
in beak sculpture is the best way to distinguish between two species. Other exterior
shell characteristics may include a prominent posterior ridge extending from beak
to posterior-ventral margin, a unique texture to the periostracum, or a wing-like
structure extending from the dorsal margin.
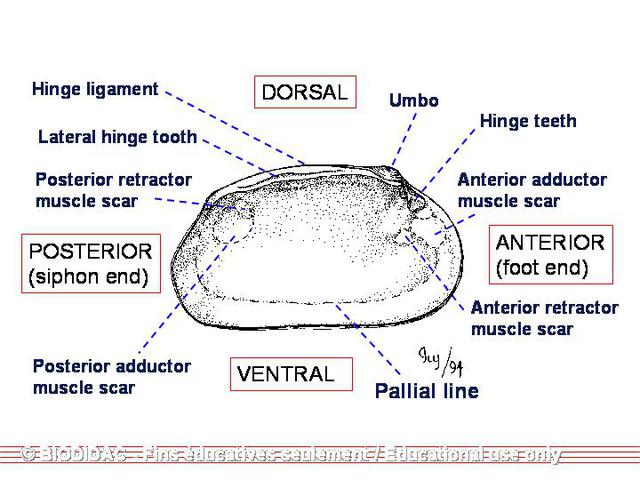
Aside from the exterior surface of the shell, researchers involved in identifying mussel species examine various aspects of the interior of the shell as well. In fact because of the high individual variability of the exterior, the interior characteristics are relied more heavily upon in identification. Probably the most important interior features are the size, shape, number, and orientation of the hinge teeth. Pseudocardinal teeth are situated slightly anterior to the beak and are generally short and triangular in shape. These teeth can vary from being broad and triangular to thin and elevated and are generally serrated. Orientation of the pseudocardinals refers to the angle between the pseudocardinals and the lateral teeth, which can be obtuse (angled away from the center of the shell), nearly right (vertical), or acute (angled toward the center of the shell). In some species, the pseudocardinal teeth are practically parallel with the lateral teeth. Lateral teeth are the long, slender, raised ridges posterior to the beak. These teeth can vary in length; can be straight or curved, smooth or serrated, thick or thin, and compressed or elevated. In some species, the teeth are well-developed, while in other species, the teeth are poorly-developed or reduced to a thickening along the hinge. In addition to the characteristics of the hinge teeth, mussels vary in the depth of the beak cavity, the width of the interdentum (the area between the pseudocardinal and lateral teeth along the hinge), the color of the nacre, the degree of impression of the muscle scars and pallial line (the line running parallel to the ventral margin), the thickness of the shell, the degree of inflation of the shell (the width or distance from one beak to the other when the two valves are together), the curvature of each margin, and the height of the umbo. Most species are identified by a combination of several characteristics.
{kind=link}
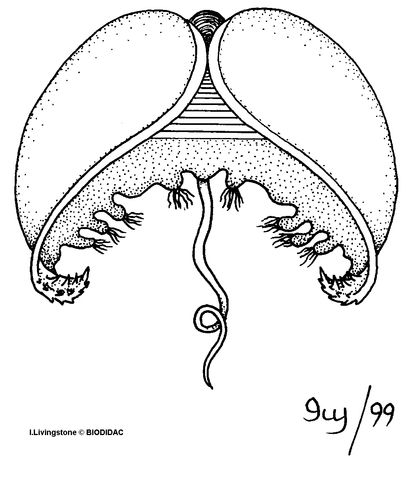
Glochidia
are the parasitic stage of the larvae and are generally dependent on a host to survive.
Mature glochidia range from 0.05 to 0.5 mm in diameter. They are bivalves, which vary
in shape from triangular, circular, oblong, or (in
Potamilus
only) ax head shaped and are typically attached by a single adductor muscle. Most
glochidia have sensory hairs lining their mantle and a larval thread protruding from
the open valves, which may allow them to attach to the host. Many species have hook-like
structures to allow them to attach to the fins or skin of the fish. Those species
without hooks usually attach to the gills.
{kind=link}
Measurements are generally taken of the length, height, and width. The length is the distance from the anterior to the posterior margin. The height is the distance from dorsal to ventral margin, usually at the beak. Width is the widest point when the mussel valves are together, which is usually below the beaks. In addition, some identification keys will use the length to height ratio as a way to distinguish some species.
- Other Physical Features
- ectothermic
- heterothermic
- bilateral symmetry
- polymorphic
- Sexual Dimorphism
- sexes alike
- sexes shaped differently
Development
Embryonic unionids develop within the marsupia, or specialized portions of the gills,
of the female. Once fully developed, they are
released
from the female and must attach to the gills or fins of a fish host within a few
days or they will die.
Strophitus undulatus
and
Utterbackia imbecillis
are the only two species capable of direct development without a host (Watters 1994c).
Only one species,
Simpsonaias ambigua
, metamorphoses on a salamander,
Necturus maculosus
. Many unionids are species-specific, requiring one or a narrow range of species.
If attached to the wrong species, the glochidia will die as a result of the fish's
immune system response (Watters 1998). Within a couple of days, the hosts’ dermal
tissue will encapsulate each glochidium forming a nodular cyst. While encysted, the
glochidia will metamorphose, allowing the organs to develop more like an adult’s organs
(Meglitsch & Schram 1991). There is a mortality rate of over 99.99% from the time
the glochidia are released from the mother to the time in which the metamorphosed
juveniles settle in the sediments (Jansen et al 2001).
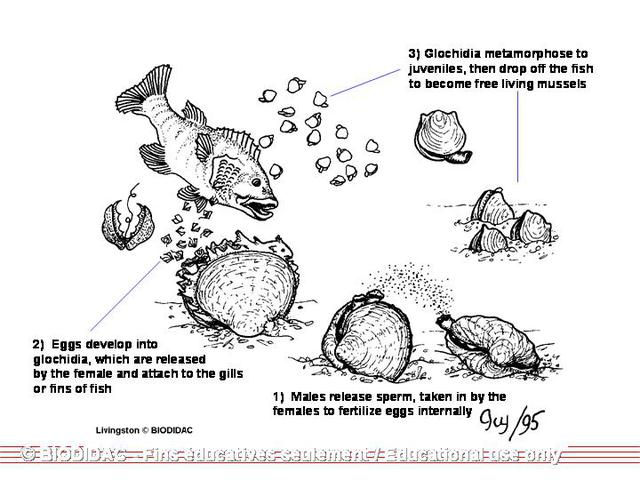{kind=link}
After an average of 10-30 days (the record is 190 days), the metamorphosis will be
complete and the glochidia will break from the cysts and drop from the host. The
third and final stage of development occurs in the sediments of the stream or lake
and may last anywhere from one to eight years before the juvenile is sexually mature.
In this juvenile stage, the young mussel will complete its internal development, create
the adult shell, and begin to live independently in the stream or lake.
As in most bivalves, the shell is composed of three layers: the periostracum, the
prismatic layer, and the nacre. The periostracum is the outermost layer and is composed
of an organic material. The prismatic layer is the middle layer and is composed of
thin blocks of a prism-like calcium carbonate, which are oriented perpendicular to
the mantle and the other two layers. The nacre, or mother of pearl, is the innermost
layer, which is composed of thin, alternating, laminae (flakes or sheets) of calcium
carbonate and an organic material (Smith 2001). The mantle is responsible for generating
new shell as the mussel ages. A mantle flap is pressed against the interior of each
valve and ends in three folds. The periostracum forms at the outer margin and the
prismatic layer forms at the outer border. The nacre forms along the entire surface
of the mantle.
Muscle scars
form where the muscle attaches to the shell, disrupting the formation of the nacre.
Instead of the shell forming along the dorsal edge where the hinge is located, an
elastic hinge ligament composed of conchiolin (a protein-rich substance) forms, binding
the two valves together (Meglitsch & Schram 1991).
{kind=link}
Growth of the mussel begins at the elevated portion called the
umbo or beak
. Because new shell is added along the entire edge of the mantle, concentric rings
form around the beak. In some species, these rings may be grouped closer together
in some areas than others, forming ridges. These ridges indicate the period of diapause
during the winter or unfavorable environmental conditions, such as lower water level
or lack of food. The period of growth in northern populations is typically from April
to September. The growth rate depends mostly on environmental conditions such as
water temperature, food supply, and the chemical composition of the water. Many mussel
species are capable of growing 30 to 80 mm every two growing seasons.
{kind=link}
- Development - Life Cycle
- metamorphosis
Reproduction
A few species are occasionally or permanently simultaneous hermaphrodites (Bauer 1987),
but in most cases, unionid sexes are separate. Bauer (1987) suggested that hermaphroditism
occurs when the population density is low or gene flow is limited. In these cases,
the female is the only one of the two sexes that can become hermaphroditic. Despite
the dioecious nature of most mussels, males and females do not make contact with each
other. Males produce sperm year round and
release
during the time of year when females ovulate (Matteson 1948). This simultaneous
release of gametes may be triggered by a change in the water temperature and the intensity
of light in the environment. The male’s sperm leaves the suprabranchial chamber of
each demibranch and exits the organism through the exhalant aperture to be carried
by the water current to a nearby female. Because sperm cannot swim against the current,
the receiving female must be downstream (Watters 1994a). The sperm enters the female
through the inhalant aperture and fertilizes the eggs stored in the water tubes of
the demibranch’s lamellae (Smith 2001).
- Mating System
- polygynandrous (promiscuous)
Depending on the species, sexual maturity is reached between one and eight years (Smith
2001). Gamete production is initiated by a change in the water temperature surrounding
the mussel (Watters 1998). Annual gametogenesis and gravidity may occur throughout
the year or during certain seasons depending on latitude. The more northern populations
tend to be gravid for a few months or all winter long and release the glochidia in
the spring. There are a few species that release the glochidia in the fall. In many
cases, southern populations are not restricted to reproducing during certain seasons.
The number of larvae developing in one female at a time may range from several thousand
in some of the smaller
Unionidae
genera to possibly over 1 million. The maximum amount of glochidia in one female
is unknown, but Tankersley and Dimock (1992) recorded nearly 1 million in a
Pyganodon cataracta
female. The closest related family,
Margaritiferidae
, contains species which have produced more than 3 million per individual (Smith 2001).
Bradytictic (long term) breeders will maintain the glochidia within the marsupia,
the specialized portions of the gills, until the following spring or summer before
releasing them into the water
. Tachytictic (short term) breeders will release the glochidia in the same year,
usually by July or August (Watters 1998). Matteson (1948) was convinced that the
membrane surrounding the developing embryos provides all of the necessary nutrients,
rather than the female transferring food to the developing young. His conclusion
was based on a lack of connective structure from the gills to the young and that the
fertilization membrane surrounding each embryo, which prevents the passing of any
materials, remains until development is complete.
{kind=link}
- Key Reproductive Features
- iteroparous
- seasonal breeding
- year-round breeding
- gonochoric/gonochoristic/dioecious (sexes separate)
- simultaneous hermaphrodite
- sexual
- fertilization
- ovoviviparous
Unionid embryos spend the first stage of development in the marsupial portion of the
female unionid's gills, where they develop into glochidia, the parasitic stage. Once
the first stage is complete, usually in the spring, the female will
release the glochidia
into the water to begin the second stage as a parasite. Because glochidial mortality
is high, many unionids have developed specialized methods of attracting fish to the
mother before the glochidia are released, increasing the chances the larvae can attach
to a host. Some of these species extend the glochidia encapsulated in conglutinates
(Chamberlain 1934). These conglutinates (sacs) are attached to the parent organism
and move in the current like worms. This encapsulated appendage acts as a lure to
attract the host fish, which then eats the glochidia freeing them from the capsule
and allowing them to attach to the gills of the fish. Other species use a modified
mantle flap to attract the fish. This flap mimics the prey of the potential host
fish. The glochidia are sensitive enough to attach themselves to the fish as soon
as contact is made.
{kind=link}
- Parental Investment
-
pre-fertilization
- provisioning
-
protecting
- female
-
pre-hatching/birth
-
provisioning
- female
-
protecting
- female
-
provisioning
Lifespan/Longevity
For small organisms, unionids are long-lived, living an average of 10 or more years
(Cummings & Mayer 1992). Some genera live only 8 to 9 years, while others can live
up to 10 to 15 years (Smith 2001). Given the proper conditions, many species can live
up to 20 or 30 years (Watters 1998). Bauer (2001b) suggested life span is dependent
upon metabolic rate. Mussels with a higher metabolic rate tend to have a shorter
life span. Those unionids in larger rivers or streams would have a higher metabolic
rate due to the abundance of food, and would be expected to have a short life. Unionids
that thrive further upstream may have a longer lifespan because they would have adapted
to a limited food supply by decreasing their metabolic rate. Although metabolic rate
is a key factor affecting longevity in some species, it is not a universal constant.
Some species with similar metabolic rates may have very different lifespans.
Behavior
For the most part, mussels are sedentary, but they are capable of a restricted form
of locomotion. They move around by a series of muscular motions of the
foot
located at the anterior end of each individual. The foot is thrust forward first.
It then swells and shortens at the same time, causing the body and shell to pull forward
slightly. This process is repeated until the mussel has reached its destination.
Some species have been recorded to move up to several feet within an hour. Researchers
are still unsure what causes this migration, but they suspect the movement is caused
by a drop in water level or some other unfavorable change in the surrounding environment.
Unionids are solitary organisms. The only intra- or inter-species interactions occur during reproduction. Once they drop from the host , the mussel becomes a solitary individual and live partially buried in the sediments. As juveniles, mussels burrow into the sediments along the bottom of the stream or lake, which protects them from predators. Once mature, more of the organism must protrude from the substrate in order for the inhalant and exhalant apertures to bring in and expel water. Because more of the shell is visible, they are more susceptible to predation.
During winter months and aestivation periods (Matteson 1955; van der Schalie 1940),
mussels will burrow into the substrate until only the apertures are protruding. They
then go into a state of dormancy where the apertures only open on occasion. Some
genera are able to survive in this dormant state among the dry to moist sediments
for months at a time (Smith 2001).
- Key Behaviors
- parasite
- motile
- sedentary
- hibernation
- solitary
Communication and Perception
Unionids use specialized structures to visually attract potential fish hosts. The
combination of the statocysts and the statolith aids the mussel in maintaining equilibrium
by sensing gravity. They may also be able to detect vibrations (Meglitsch & Schram
1991). Although the function of the osphradia is uncertain, some researchers believe
that they detect foreign particles brought in through the inhalant aperture (Smith
2001). Drastic changes in the intensity of the light in the environment can be detected
by the mantle border (Smith 2001). Glochidia can usually detect light changes with
ocelli, but the eyes are generally lost after metamorphosis (Meglitsch & Schram 1991).
Many mussel species also have tactile cells lining the exposed portion of the mantle,
which aid in the organism's sense of touch (Meglitsch & Schram 1991). The glochidia
are especially sensitive to touch, which helps in the attachment to a host as it comes
close to them (Arey 1921).
- Other Communication Modes
- mimicry
- Perception Channels
- visual
- tactile
- vibrations
- chemical
Food Habits
Adult freshwater mussels are filter feeders; they continuously filter food particles
out of the water (Watters 1998; Allen 1921). Water is constantly pumped into the
inhalant aperture
, through the gills, and out the exhalant aperture by cilia. The cilia lining the
inner surface of the mantle, demibranchs, and visceral mass create a current by beating
in a coordinated manner. Organic and inorganic particles suspended in the water surrounding
the inhalant aperture are brought in by the current and caught in the mucus lining
the demibranchs. The constant current created by the cilia moves the mucus with any
trapped particles to the cilia lining the
labial palps
. The labial palps remove the inorganic particles and push them toward the ventral
margin where they drop off, are move by the cilia backward, and released between the
valves just below the inhalant aperture (Smith 2001). The organic particles are separated
by size in sorting areas on the labial palps and are then directed into the mouth.
From the mouth, particles are moved through a short esophagus to the
digestive gland
surrounding the stomach. Food particles enter the stomach through the subdivided
pores of the large digestive gland (Meglitsch & Schram 1991). Small particles are
digested intracellularly as they enter the stomach. The intestinal glands are responsible
for phagocytosis, intracellular digestion, food absorption, secretion of enzymes and
excretion (Meglitsch & Schram 1991). The intestine coils behind and below the stomach
before it extends dorsally and empties into the mantle cavity through the anus located
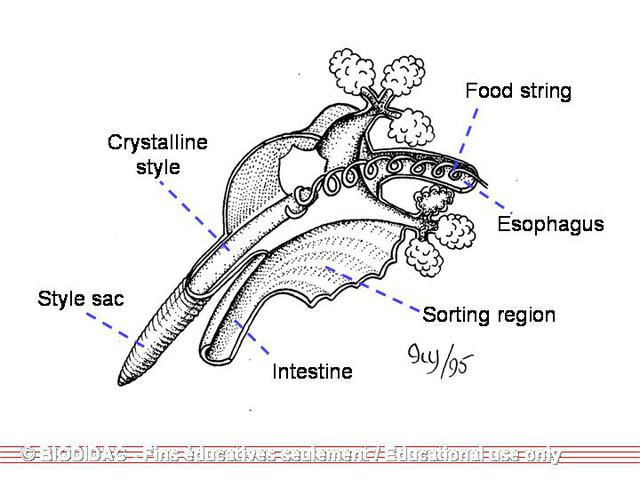
just above the exhalant aperture. At the anterior end of the stomach is a lateral
diverticulum (groove) containing a
crystalline style
(an elongated structure composed of a dense gelatinous material) (Smith 2001). The
style extends into the stomach and rotates clockwise by the ciliary epithelium within
the style sac (Meglitsch & Schram 1991; Smith 2001). The anterior end, which is in
the stomach, erodes as it rubs against the digestive shield. As is erodes, it releases
the digestive enzymes, amylase, lipase, and cellulase (Meglitsch & Schram 1991). Because
unionids do not ingest large particles, the ciliary action surrounding the style returns
the larger particles back to the entrance of the digestive glands to be resorted.
During periods of starvation, the crystalline style will disappear, but will regenerate
when food is more abundant (Smith 2001). Regeneration is a slow process which occurs
during low water temperatures.
{kind=link}
{kind=link}
The exact type of food consumed by adult freshwater mussels has been debated for some
time now. Some researchers have suggested mussels eat algae and diatoms (Allen 1914),
while others suggest bacteria, protozoans and other organic particles were ingested
(Watters 1998). A few studies have even suggested ingesting silt somehow enhances
the survival of the organism (Watters 1998). Generally, unionids feed on the bacteria
and microphytoplankton but nothing larger (Smith 2001; Cummings & Mayer 1992).
The phagocytic mantle cells of the glochidia feed off of the host’s tissue (Meglitsch
& Schram 1991). Before attachment, glochidia must locate a proper host. In most cases,
they end up in the stream or lake sediments with the open end of the valves up awaiting
a fish to brush up against the mud allowing the larvae to attach themselves to the
fins. The glochidia of other species swim around in the water by clapping the valves
together.
- Primary Diet
- planktivore
- detritivore
- Foraging Behavior
- filter-feeding
Predation
Muskrats
are probably the most important mammal that preys on freshwater mussels (Cummings
& Mayer 1992; Smith 2001). These animals drag the mussels, primarily the thin-shelled
species, on the shore and either break the shells open with their teeth or leave them
on the banks until the mussel dies and the shell opens (Smith 2001). In active muskrat
foraging areas, there are often middens of a variety of shells which have been cleaned
by the muskrats. Other common unionid predators include
minks
,
otters
,
raccoons
,
turtles
,
hellbenders
, fish, some species of birds, and humans (Cummings & Mayer 1992; Smith 2001; Watters
1998). Some of the common fish species include the
freshwater drum
,
sheepshead
,
lake sturgeon
,
spotted suckers
,
redhorses
, and
pumpkinseed
.
Shortnose sturgeon
often consume
Elliptio
juveniles.
Catfish
and sheepshead have been found preying upon the more soft-shelled species (Smith
2001). In Europe,
hooded crows
have been known to prey upon mussels. They are able to reach the soft tissue by
dropping the mussels to crack the shell open (Watters 1998).
Aside from predators, mussels are often invaded by destructive parasites.
Unionicolid mites
and
monogenean trematodes
are often found feeding upon the mantle and branchial tissue. A chironomid midge,
Baeoctenus bicolor
, feeds and lays its eggs upon the demibranchs (Smith 2001) and has been known to
consume up to 50% of the gill tissues (Watters 1998), which interferes with the respiration
and reproduction of the mussel. Other trematodes infect the nacre causing the formation
of irregular pearls and blisters and often discolor the nacre (Smith 2001).
Aspidogastrean trematodes
are known to infect the branchial, intestinal, and pericardinal cavities (Smith 2001).
Matteson (1955) found a
leech
attached to the mantle cavity of a female
Lampsilis siliquoidia
.
To avoid these predators, mussels will bury themselves into the lake or stream sediments.
Because adult unionids do not have true siphons, only openings in the mantle, they
must leave the posterior margin out of the sediments to allow for sufficient respiration.
This exposure leaves the organism vulnerable to predation, desiccation, and temperature
extremes (Watters 1998).
Ecosystem Roles
Like all other organisms, freshwater mussels play an important role within their ecosystem.
Not only do they provide a food source for muskrats and other predators, but they
also aid in the decomposition of detritus and keep the bacterial and planktonic populations
under control (Pusch et al 2001; Jorgensen 1990). Dense mussel populations rely on
rapid currents for survival. During periods of little or no current, these dense
mussel beds can cause a depletion of the dissolved oxygen and food supply, causing
a rise in the mortality rate of the mussel and other faunal populations along the
basin (Jorgensen 1990).
Researchers have found that the glochidia generally do not cause sufficient enough
damage to the host to cause problems. Cases of over 3000 glochidia infecting a fish
without apparent harm have been reported. However, there have also been cases where
30 mm fingerling trout have died of secondary bacterial infections caused by a little
more than 100 glochidia (Smith 2001). Some fish species are able to develop an immune
response to resist the glochidia causing them to pre-maturely drop off the fish.
- Ecosystem Impact
- parasite
- large-mouth bass Micropterus salmoides
- long-nosed gar Lepisosteus osseus
- orange spotted sunfish Lepomis humilis
- bitterling Rhodeus sericeus
- yellow perch Perca flavescens
- walleye Stizostedion vitreum
- white-tail shiner Cyprinella galacturus
- spotfin shiner Cyprinella spilopterus
- big-eye chub Hybopsis amblops
- mud puppy Necturus maculuosus
- aspidogastrean trematodes ( Aspidogastrea )
- monogenean trematodes Monogenea
- unionicolid mites Unionicolidae
- a chironomid midge Baeoctenus bicolor
- leeches Hirudinea
Economic Importance for Humans: Positive
Humans have used freshwater mussels and their shells for food, jewelry, tools, utensils,
and pottery temper for thousands of years (Cummings & Mayer 1992). Native Americans
have been carving shells for implements and ornamentation for at least 3000 years.
Around 1000 years ago, people in North America discovered that tempering their pottery
with crushed shells rather than sand or gravel allowed them to create a smoother,
thinner vessel. During this same period, people were creating beads, hoes and spoons
with freshwater mussel shells (Wiant 2000).
Before 1890, freshwater mussels were used for only a few decorative items such as
pistol grips, brush handles, and jewelry. Both U.S. tariffs on imported goods (including
buttons) and the rise of the new ready-to-wear clothing industry created high demand
for buttons. The pearl button industry began in 1891 with the start of a new fashion
trend to use shell buttons to fasten clothes. With Muscatine, Iowa as the center of
the industry, pearl button production became a major industry for hundreds of river
towns along the Mississippi and other Midwestern rivers. Thousands of tons of mussels
were harvested. The demand was so high that by 1900 the Illinois and Wabash rivers
were depleted of mussels. The peak of the industry occurred in 1909 with a record
of 2600 boats on the Mississippi River alone. By the 1940s and 1950s, cheaper plastic
buttons became widely available and replaced the shell buttons, causing the collapse
of this industry and the recovery of many depleted mussel populations.
In the 1950s, the Japanese pearl growers developed another use for freshwater mussel
shells (Cummings & Mayer 1992). They discovered that small beads could be carved out
of the shells of freshwater mussels and inserted into oysters to artificially form
pearls. They also found that certain species, particularly the
threeridge
and the
washboard
, were easier to manipulate and created a better nucleus. These discoveries were
the beginning of the cultured pearl industry. Today, thousands of tons of freshwater
mussel shells from North America are exported to Japan to support the pearl industry
(Cummings & Mayer 1992).
In addition to the many products, freshwater mussels act as water quality indicators. Because they are filter-feeders, pollutants in the water will accumulate in the tissue of mussels until they reach a toxic level killing the organism. A drastic drop in the mussel population is an indication of poor water quality.
- Positive Impacts
- food
- body parts are source of valuable material
- research and education
Economic Importance for Humans: Negative
There are no known adverse effects of unionid mussels on humans.
Conservation Status
Worldwide, freshwater mussels are one of the most endangered groups with significant
population declines documented in recent surveys. In the United States alone, nearly
70 species are either endangered or threatened (USFWS 2003). Reasons for the past
decline include the effects of the pearl button industry of the late 19th and early
20th centuries and the cultured pearl industry of the past 50 years. Today, siltation
from agriculture, forestry, and construction smothers the organisms inhibiting feeding
and respiration. Impoundments alter the habitat, killing first the mussels that thrive
in rapid currents. Dams cause an increase in silt as well as a constant cold water
temperature. Since many mussel species are temperature sensitive, the cold will slow
the growth and may inhibit the reproduction of the mussels that survived the initial
shock of the construction. In-stream sand and gravel mining often buries, crushes,
or removes the mussels in the substrate and releases silt, which affects the species
downstream. Agricultural runoff is another threat to mussel populations. Many species
cannot tolerate pollutants introduced in the water from pesticides, herbicides, and
fertilizers. At sub-lethal concentrations these chemicals inhibit respiration and
accumulate in the tissues of the organism. Mussels are also sensitive to heavy metals
which accumulate in the tissues. Mine runoff creates an acidic pH in the water, which
many mussel species cannot tolerate for long periods of time. Salinity from road
salt runoff is lethal to glochidia.
In addition to industrial wastes and depletion, mussels now compete for resources with introduced species. The Asian clam and the zebra mussel are probably the two most common exotic species, which have been introduced to North American freshwaters.
Other Comments
The study of freshwater mussels as a group is made more complicated by a particularly
high rate of duplicate species names. Many 19th century biologists didn't realize
that members of the same mussel species grow somewhat differently-shaped shells depending
on the conditions of the stream or lake they are growing in. The biologists described
new species based on shell shape, and consequently there are many hundreds of duplicate
names for the same species.
Additional Links
Contributors
Lisa Winhold (author), Animal Diversity Web.
Renee Sherman Mulcrone (editor).
- Nearctic
-
living in the Nearctic biogeographic province, the northern part of the New World. This includes Greenland, the Canadian Arctic islands, and all of the North American as far south as the highlands of central Mexico.

- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Palearctic
-
living in the northern part of the Old World. In otherwords, Europe and Asia and northern Africa.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- oriental
-
found in the oriental region of the world. In other words, India and southeast Asia.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- Australian
-
Living in Australia, New Zealand, Tasmania, New Guinea and associated islands.
- native range
-
the area in which the animal is naturally found, the region in which it is endemic.
- holarctic
-
a distribution that more or less circles the Arctic, so occurring in both the Nearctic and Palearctic biogeographic regions.
Found in northern North America and northern Europe or Asia.
- temperate
-
that region of the Earth between 23.5 degrees North and 60 degrees North (between the Tropic of Cancer and the Arctic Circle) and between 23.5 degrees South and 60 degrees South (between the Tropic of Capricorn and the Antarctic Circle).
- tropical
-
the region of the earth that surrounds the equator, from 23.5 degrees north to 23.5 degrees south.
- freshwater
-
mainly lives in water that is not salty.
- benthic
-
Referring to an animal that lives on or near the bottom of a body of water. Also an aquatic biome consisting of the ocean bottom below the pelagic and coastal zones. Bottom habitats in the very deepest oceans (below 9000 m) are sometimes referred to as the abyssal zone. see also oceanic vent.
- intertidal or littoral
-
the area of shoreline influenced mainly by the tides, between the highest and lowest reaches of the tide. An aquatic habitat.
- ectothermic
-
animals which must use heat acquired from the environment and behavioral adaptations to regulate body temperature
- heterothermic
-
having a body temperature that fluctuates with that of the immediate environment; having no mechanism or a poorly developed mechanism for regulating internal body temperature.
- bilateral symmetry
-
having body symmetry such that the animal can be divided in one plane into two mirror-image halves. Animals with bilateral symmetry have dorsal and ventral sides, as well as anterior and posterior ends. Synapomorphy of the Bilateria.
- polymorphic
-
"many forms." A species is polymorphic if its individuals can be divided into two or more easily recognized groups, based on structure, color, or other similar characteristics. The term only applies when the distinct groups can be found in the same area; graded or clinal variation throughout the range of a species (e.g. a north-to-south decrease in size) is not polymorphism. Polymorphic characteristics may be inherited because the differences have a genetic basis, or they may be the result of environmental influences. We do not consider sexual differences (i.e. sexual dimorphism), seasonal changes (e.g. change in fur color), or age-related changes to be polymorphic. Polymorphism in a local population can be an adaptation to prevent density-dependent predation, where predators preferentially prey on the most common morph.
- metamorphosis
-
A large change in the shape or structure of an animal that happens as the animal grows. In insects, "incomplete metamorphosis" is when young animals are similar to adults and change gradually into the adult form, and "complete metamorphosis" is when there is a profound change between larval and adult forms. Butterflies have complete metamorphosis, grasshoppers have incomplete metamorphosis.
- polygynandrous
-
the kind of polygamy in which a female pairs with several males, each of which also pairs with several different females.
- iteroparous
-
offspring are produced in more than one group (litters, clutches, etc.) and across multiple seasons (or other periods hospitable to reproduction). Iteroparous animals must, by definition, survive over multiple seasons (or periodic condition changes).
- seasonal breeding
-
breeding is confined to a particular season
- year-round breeding
-
breeding takes place throughout the year
- sexual
-
reproduction that includes combining the genetic contribution of two individuals, a male and a female
- fertilization
-
union of egg and spermatozoan
- internal fertilization
-
fertilization takes place within the female's body
- ovoviviparous
-
reproduction in which eggs develop within the maternal body without additional nourishment from the parent and hatch within the parent or immediately after laying.
- parasite
-
an organism that obtains nutrients from other organisms in a harmful way that doesn't cause immediate death
- motile
-
having the capacity to move from one place to another.
- sedentary
-
remains in the same area
- hibernation
-
the state that some animals enter during winter in which normal physiological processes are significantly reduced, thus lowering the animal's energy requirements. The act or condition of passing winter in a torpid or resting state, typically involving the abandonment of homoiothermy in mammals.
- solitary
-
lives alone
- visual
-
uses sight to communicate
- chemical
-
uses smells or other chemicals to communicate
- mimicry
-
imitates a communication signal or appearance of another kind of organism
- visual
-
uses sight to communicate
- tactile
-
uses touch to communicate
- vibrations
-
movements of a hard surface that are produced by animals as signals to others
- chemical
-
uses smells or other chemicals to communicate
- filter-feeding
-
a method of feeding where small food particles are filtered from the surrounding water by various mechanisms. Used mainly by aquatic invertebrates, especially plankton, but also by baleen whales.
- parasite
-
an organism that obtains nutrients from other organisms in a harmful way that doesn't cause immediate death
- food
-
A substance that provides both nutrients and energy to a living thing.
- planktivore
-
an animal that mainly eats plankton
- detritivore
-
an animal that mainly eats decomposed plants and/or animals
References
Allen, W. 1921. Studies of the Biology of Freshwater Mussels: Experimental Studies of the Food Relations of Certain Unionidae. Biological Bulletin , 40: 210-241.
Allen, W. 1914. The Food and Feeding Habits of Freshwater Mussels. Biological Bulletin , 27: 127-146.
Arey, L. 1921. An Experimental Study on Glochidia and the Factors Underlying Encystment. Journal of Experimental Zoology , 33: 463-499.
Bauer, G. 2001a. Characterization of the Unionoida (=Naiads). Pp. 3-4 in Ecological Studies: Ecology and Evolution of the Freshwater Mussels Unionoida , Vol. 145. Berlin: Springer-Verlag.
Bauer, G. 2001b. Framework and Driving Forces for the Evolution of Naiad Life Histories. Pp. 233-255 in Ecological Studies: Ecology and Evolution of the Freshwater Mussels Unionoida , Vol. 145. Berlin: Springer-Verlag.
Bauer, G. 1987. Reproductive Strategy of the Freshwater Pearl Mussel Margaritifera margaritifera. Journal of Animal Ecology , 56: 691-704.
Burch, J. 1975. Freshwater Unionacean Clams (Mollusca: Pelecypoda) of North America . Ann Arbor, MI: University of Michigan.
Chamberlain, T. 1934. The Glochidial Conglutinates of the Arkansas Fanshell, Cyprogenia aberti (Conrad). Biological Bulletin , 66: 55-61.
Cummings, K., C. Mayer. 1992. Field Guide to Freshwater Mussels of the Midwest, Manual 5 . Champaign, Illinois: Illinois Natural History Survey. Accessed July 13, 2003 at http://www.inhs.uiuc.edu/cbd/collections/mollusk/fieldguide.html .
Graf, D. 2002. Molecular phylogenetic analysis of two problematic freshwater mussel genera (Unio and Gonoidea) and a re-evaluation of the classification of Nearctic Unionidae (Bivalvia: Palaeoheterodonta: Unionoida). Journal of Molluscan Studies , 68: 65-71.
Graf, D., K. Cummings. 2008. "The Mussel Project: A Resource for Freshwater Malacologists" (On-line). Accessed December 15, 2008 at http://bama.ua.edu/~musselp/index.html .
Haas, F. 1969. Das Tierreich Lieferung 88: Superfamilia Unionacea . Berlin: Walter de Gruyter.
Huitt, S., R. Warren. 2003. "Freshwater Mussels" (On-line). Harvesting the River. Accessed August 03, 2003 at http://www.museum.state.il.us/RiverWeb/harvesting/harvest/mussels/index.html .
Jansen, W., G. Bauer, E. Zahner-Meike. 2001. Glochidial Mortality in Freshwater Mussels. Pp. 185-211 in Ecological Studies: Ecology and Evolution of the Freshwater Mussels Unionoida , Vol. 145. Berlin: Springer-Verlag.
Jennings, S. 2000. "Needs in the Management of Freshwater Mussels in the National Park System" (On-line). Accessed July 18, 2003 at http://www.nature.nps.gov/wrd/mussels/mussels5-01.htm .
Jorgensen, C. 1990. Bivalve Filter Feeding: Hydrodynamics, Bioenergetics, Physiology, and Ecology . Denmark: Olsen and Olsen.
Kohl, M. 2000. "Unionidae" (On-line). Freshwater Molluscan Shells. Accessed August 31, 2003 at http://members.aol.com/savetheclams/Unionidae.html .
Matteson, M. 1948. Life History of Elliptio complanatus (Dillwyn, 1817). American Midland Naturalist , 40: 690-723.
Matteson, M. 1955. Studies on the Natural History of the Unionidae. American Midland Naturalist , 53: 126-145.
Meglitsch, P., F. Schram. 1991. Invertebrate Zoology, Third Edition . New York, NY: Oxford University Press, Inc.
Pusch, M., J. Siefert, N. Walz. 2001. Filtration and Respiration Rates of Two Unionid Species and Their Impact on the Water Quality of a Lowland River. Pp. 317-326 in Ecological Studies: Ecology and Evolution of the Freshwater Mussels Unionoida , Vol. 145. Berlin: Springer-Verlag.
Reigle, N. 1967. An occurrence of Anodonta (Mollusca, Pelecypoda) in Deep Water. American Midland Naturalist , 78: 530-531.
Salmon, A., R. Green. 1982. Environmental Determinants of Unionid Clam Distribution in the Middle Thames River. Canadian Journal of Zoology , 61: 832-838.
Simpson, C. 1914. A Descriptive Catalogue of the Naiads, or Pearly Fresh-water Mussels, Parts I-III . Detroit, MI: Bryant Walker.
Simpson, C. 1900. Synopsis of the Naiads, or Pearly Fresh-water Mussels. Proceedings of the United States National Museum, 22: 501-1044.
Smith, D. 2001. Pennak's Freshwater Invertebrates of the United States: Porifera to Crustacea, Fourth Edition . New York: John Wiley & Sons, Inc..
Strayer, D., J. Ralley. 1993. Microhabitat use by an assemblage of stream-dwelling unionaceans (Bivalvia), including two rare species of Alasmidonta. Journal of the North American Benthological Society , 12: 247-258.
Strayer, D. 1983. The effects of surface geology and stream size on freshwater mussel (Bivalvia: Unionidae) distribution in southeastern Michigan, U. S. A.. Freshwater Biology , 13: 253-264.
Tankersley, R., R. Dimock Jr. 1992. Quantative Analysis of the Structure and Function of the Marsupial Gills of the Freshwater Mussel Anodonta cataracta . Biological Bulletin , 182: 145-154.
Turgeon, D., J. Quinn, A. Bogan, E. Coan, F. Hochberg, W. Lyons, P. Mikkelsen, R. Neves, C. Roper, G. Rosenberg, B. Roth, A. Scheltema, F. Thompson, M. Vecchione, J. Williams. 1998. Common and Scientific Names of Aquatic Invertebrates from the United States and Canada: Mollusks, Second Edition . Bethesda, Maryland: American Fisheries Society, Special Publication 26.
U.S. Fish and Wildlife Service, 2003. "Species Information: Threatened and Endangered Animals and Plants" (On-line). Accessed August 13, 2003 at http://endangered.fws.gov/wildlife.html#Species .
Warren, R. 2000. "Native Americans: Historic" (On-line). Museum Link Illinois. Accessed August 03, 2003 at http://www.museum.state.il.us/muslink/nat_amer/post/index.html .
Watters, G. 1994c. An Annotated Bibliography of the Reproduction and Propagation of the Unionoidea (Primarily of North America) . Columbus, Ohio: Ohio Biological Survey: Miscellaneous Contributions No. 1.
Watters, G. 2001a. "Electric Elliptio Land in North America" (On-line). Accessed July 18, 2003 at http://www.biosci.ohio-state.edu/~molluscs/Elliptio/ .
Watters, G. 1998. "Freshwater Mussels: Biology" (On-line). Conchologists of America Conch-net. Accessed July 25, 2003 at http://coa.acnatsci.org/conchnet/uniobio.html .
Watters, G. 1994a. North American Freshwater Mussels Part 1: The Quick and the Dead. American Conchologist , 22: 4-7. Accessed December 15, 2008 at http://www.conchologistsofamerica.org/articles/y1994/9406_watters.asp .
Watters, G. 1994b. North American Freshwater Mussels Part 2: Identification, Collection, and the Art of Zen Malacology. American Conchologist , 22: 11-13, 18. Accessed December 15, 2008 at http://www.conchologistsofamerica.org/articles/y1994/9409_watters.asp .
Watters, G. 2001b. The Evolution of the Unionacea in North America, and Its Implications for the Worldwide Fauna. Pp. 281-307 in Ecological Studies: Ecology and Evolution of the Freshwater Mussels Unionoida , Vol. 145. Berlin: Springer-Verlag.
Watters, G. 1992. Unionids, Fishes, and the Species-Area Curve. Journal of Biogeography , 19: 491-490.
Wiant, M. 2000. "Native Americans: Prehistoric" (On-line). Museum Link Illinois. Accessed August 03, 2003 at http://www.museum.state.il.us/muslink/nat_amer/pre/index.html .
van der Schalie, H. 1940. Aestivation of Fresh-Water Mussels. Nautilus , 43: 137-138.